目录
更新于 – 2026 年 4 月 28 日
ADHD/ADD标准疗法与芳香疗法(使用治疗级、经GC/MS分析批次检测的精油)的比较研究。(截至2026年4月)。.
ADHS/ADS 与精油——通俗易懂的解释
一份面向所有想了解天然ADHD辅助疗法的人的、通俗易懂的科学报告简述
什么是ADHD/ADD?
ADHD 代表 注意力缺陷多动障碍, 广告 对于没有多动症(注意力缺陷障碍)的版本。两者都不是虚构的或由父母不当教养造成的,它们是真实存在的神经系统疾病,大脑的连接方式与常人不同。.
后面的图片: 想象一下大脑就像一台收音机。多动症的大脑,它的频道一直在不同频率之间切换,有时声音太大,有时声音太小,很少能调到正确的频率。这使得人们很难集中注意力、静坐或控制冲动。.
ADHD 大脑中会发生什么?
核心问题在于两种神经递质:
多巴胺 “奖赏和激励的信使”
在ADHD中,多巴胺被(通过多巴胺转运体DAT)过快地“清理”掉。这导致日常任务缺乏奖励感和动力。大脑不断寻求刺激,因此会产生冲动和冒险行为。
去甲肾上腺素 “注意力与专注信使”
过少的去甲肾上腺素在大脑“指挥中心”——前额叶皮层中——导致注意力不集中、健忘、计划困难
哪些大脑区域受到影响?
- 前额叶皮层
负责计划、冲动控制、专注——ADHD 时活动减弱
- 纹状体(基底神经节)
奖赏中枢——多动症患者敏感度较低
- 小脑
时机与协调——ADHD 患者略有不同
ADHD 的患病率是多少?
根据儿童和青少年,这是最常见的精神疾病,其中 5 到 7 %, 在成人中 2.5 至 4 %, 许多成年人不知道自己患有ADHD。.
ADHD 在儿童和成人中的表现方式不同。**儿童 ADHD 的迹象和症状可能包括:*** **注意力不集中:** * 难以集中注意力或长时间保持注意力 * 容易分心 * 难以跟随指示或完成任务 * 经常丢东西 * 健忘 * 避免需要持续脑力活动的任务 * **冲动:** * 冲动地说话或插话 * 难以等待自己的回合 * 在不恰当的时候行动或说话 * 难以控制情绪 * **多动:** * 坐立不安或难以保持静止 * 不停地跑动或攀爬 * 说话过多 * 难以进行安静的活动**成人 ADHD 的迹象和症状可能与儿童有所不同,并且可能包括:*** **组织和时间管理困难:** * 难以规划和组织任务 * 拖延 * 难以按时完成截止日期 * 难以管理财务 * **情绪调节困难:** * 易怒 * 沮丧 * 情绪波动 * 难以控制愤怒 * **关系困难:** * 对他人的想法或感受不敏感 * 难以维持稳定的人际关系 * **冲动行为:** * 鲁莽驾驶 * 冲动消费 * 滥用药物 * 难以做出决定 * **注意力不集中:** * 尽管有极大的努力,但仍然难以集中注意力 * 粗心大意 * 容易分心 * 遗忘约会或承诺重要的是要注意,ADHD 的症状因人而异,并且可能随着年龄的增长而改变。如果您认为自己或您的孩子可能有 ADHD,请务必咨询医疗保健专业人员以获得准确的诊断和适当的治疗。
| 区域 | 典型症状 |
| 请注意 | 分心,粗心犯错,忘记日常事物 |
| 多动症 | 坐立不安,站起来,内心烦躁(尤其是在成年人身上) |
| 冲动 | 打断,即时决定,不耐烦 |
| 执行功能 | 计划不周,拖延症,时间盲 |
| 情绪 | 易怒,不耐烦,情绪失调 |
ADHD通常如何治疗?
主要药物——甲基苯丙胺(利他林、专注达、美迪凯)
哌甲酯(MPH)是最常开的ADHD药物,也是一线治疗方法。.
它是如何工作的?
哌甲酯阻断多巴胺转运蛋白(DAT),即多巴胺的“清除泵”。当清除的多巴胺减少时,神经细胞之间的多巴胺会增多,大脑就能更好地集中注意力。.
背后的数字
20毫克的剂量大约可以阻断. 54 % 大脑的多巴胺清除泵,40毫克大约阻断. 72 %.
其作用在30–90分钟后起效,根据制剂不同,持续4–12小时。.
它有多有效?
与 70-80 % 患者接受甲基苯丙胺(哌甲酯)的临床疗效显著,注意力、冲动控制和学业成绩的改善均有充分证据支持。.
措辞
- 速效 (例如,利他林):4-6小时,每天需服用数次
- 迟钝 (例如:Concerta、Medikinet retard):每日一次,每次8-12小时
副作用
减食欲(最常见副作用)
– 睡眠障碍(如果服用过晚)
头痛,胃部不适(通常在初期)
- 心率和血压略有升高
情绪波动、易怒(罕见)
其他药物
阿托西汀(择思达)
- 无刺激物,抑制去甲肾上腺素转运体 (NET)
– 对ADHD也有效,没有成瘾风险
– 需要 4–6 周才能达到全部效果
– 适用于同时患有焦虑症或抽动障碍的患者
苯丙胺(在德国不太常见)
– 赖右苯丙胺 (Vyvanse):自 2013 年起在德国获准用于成人
– 比哌甲酯作用更强、更持久
关因子 (Intuniv)
– 适用于儿童和青少年,当兴奋剂不适用时
作用于前额叶皮层中的去甲肾上腺素受体
ADHD 注意力缺陷多动障碍中的精油——这可能如何起作用?
精油可以通过多种方式支持ADHD大脑:
- 关于嗅觉进入大脑
香气分子激活边缘系统(情绪、动机)和前额叶皮层(注意力)——直接且快速 - 关于神经递质系统
某些萜烯会影响多巴胺、去甲肾上腺素、血清素和 GABA - 与哌甲酯互补
哌甲酯作用于DAT/NET - 精油作用于其他系统(GABA-A、CB2、TRPM8、SERT),因此具有潜在的补充作用
重要限制
关于ADHD使用精油的研究仍处于初级阶段。大多数研究是在健康的成年人身上进行的,而不是ADHD患者。直接的临床证据仍然有限。.
哪些精油可以帮助治疗注意力缺陷多动障碍 (ADHD)?
薄荷——专注力精油第一名
薄荷 (薄荷) 提供了最强有力的证据来证明注意力的改善。.
- 里面是什么?
薄荷醇(30–50 %)
薄荷脑 (10–30 %)
1,8-桉叶油 (5–10 %) - 薄荷醇有什么作用?
激活TRPM8受体——神经系统中的“冷感受器”,激活大脑
抑制由酶催化的神经递质乙酰胆碱的降解
调节GABA-A受体(轻度镇静,当过度激活时) - 临床试验(n=24)
100 微升(胶囊化的)薄荷油在 RVIP(快速视觉信息处理)测试中显著提高了持续注意力(1-3 小时后);减轻了疲劳并提高了心算能力 - 特别专题
薄荷促进 聚焦 注意力(与薰衣草相反,薰衣草倾向于分散注意力) - 如何使用
学习或工作单元前,在手帕上滴2滴;在学习室使用扩散器。
迷迭香——健忘与清醒
迷迭香 (迷迭香自古以来就被称为“健忘草”,并非浪得虚名。.
- 里面是什么?
1,8-桉树脑 (20–50 %)
樟脑 (10–20 %)
α-蒎烯(10–15 %) - 1,8-桉树脑有什么作用?
抑制乙酰胆碱酯酶,这种酶会分解神经递质(类同于阿尔茨海默氏症药物!)
激活神经系统,提高警觉性和清醒度
抗炎和抗氧化在大脑 - 临床试验 (n=144)
迷迭香香薰疗法显著改善了总体记忆和次级记忆,优于薰衣草组和对照组 - 用于ADHD
对记忆问题、“脑雾”和晨间疲劳特别有帮助 - 如何使用
早晨在香薰扩散器中使用(5滴),考试或重要任务前吸入 - 请注意: 不要在晚上使用,它会过于兴奋;鉴于其樟脑含量,癫痫患者请特别小心!
薰衣草——对抗焦虑和睡眠问题
薰衣草 (狭叶薰衣草) 是治疗ADHD伴随的焦虑和睡眠问题的最佳精油。.
- 里面是什么?
芳樟醇(25–45 %)
芳樟醇乙酸酯(25–50 %) - 芳樟醇有什么作用?
激活 GABAA 受体,即大脑的“刹车”
调节血清素、多巴胺和去甲肾上腺素
降低皮质醇(压力荷尔蒙)% 24-38% - 在ADHD(注意力缺陷多动障碍)中特别有用的
睡眠困难(ADHD 非常普遍)
伴随性焦虑障碍(约占ADHD患者的50 %)
情绪失调和易怒 - 重要提示
薰衣草可以 短期损害工作记忆, 因此,不要在学习阶段使用,而仅在晚上或用于有针对性的放松时使用 - 研究
Silexan (80 毫克薰衣草油,每日胶囊剂) 在治疗焦虑症方面效果与劳拉西泮相当。 - 如何使用
晚上在扩散器中(5滴),2滴滴在枕头上
雪松木—— theming and focus
雪松木 (阿特拉斯雪松 / 弗吉尼亚刺柏) 传统上用于治疗注意力缺陷多动障碍。.
- 里面是什么?
α-雪松烯
β-雪松酮
雪松醇 - 它的作用
改善大脑供氧;镇静;激活副交感神经 - ADHS特异性
Godfrey (2001) 报告了 ADHD 儿童的积极效果;;
雪松醇在研究中降低了心率和呼吸频率 - 如何使用
在扩散器中,用于平静的活动;与薰衣草搭配效果良好
香根草——扎根的香气
香根草 (香根草) 拥有一种深沉、泥土般的气味,常被形容为“抚慰躁动不安的灵魂”。.
- 它的作用
抗焦虑(缓解焦虑);镇静神经系统;改善睡眠 - ADHS特异性
Springer 等人(2018 年)报道称,ADHD 儿童经过每天 3 次,持续 30 天的吸入治疗后有所改善。 - 为谁
尤其是在多动性注意力缺陷多动障碍(ADHD)且伴有内心不安的版本中
肉桂——唯一一项直接的ADHD研究
肉桂 (肉桂) 拥有独特的证据:是唯一一项直接针对ADHD儿童进行的临床研究。.
- 里面是什么?
肉桂醛 (60–75 %)
丁香酚 (5–10 %) - ADHD研究(Chen等人,2008)
康复+肉桂芳香疗法(% 次肉桂鼻吸)联合治疗 6 个月,在 SNAP-IV 问卷评分上显示出比单独康复(p < 0.05)有显著更好的结果。 - 机制
肉桂醛代谢物可能具有多巴胺能作用 - 限制
这项研究存在方法学上的缺陷(样本量小,盲法不明确) - 如何使用
在扩散器中少量使用,与其他精油混合
黑胡椒 (β-石竹烯) – 针对神经炎症
β-石竹烯 由黑胡椒、薰衣草和其他植物制成:
- 激活 CB2 類ен受體 无精神活性作用
- 减轻神经炎症(大脑炎症)
- ADHD 伴有炎症迹象或情绪问题很有趣
- 易于与其他精油混合
油品概览——何时使用哪种油?
| 症状 | 推荐油 | 应用时间 |
| 注意力不集中 | 薄荷,迷迭香 | 晨间,在学习阶段之前 |
| 记忆力减退 | 迷迭香 | 早上,在任务之前 |
| 睡眠问题 | 薰衣草,雪松木,香根草 | 晚上 |
| 烦躁不安/多动 | 香根草,雪松,薰衣草 | 下午,晚上 |
| 焦虑与烦躁 | 薰衣草,佛手柑 | 下午,晚上 |
| 晨练 | 薄荷,迷迭香 | 早上 |
| ADHD 综合支持 | 肉桂(少量) | 学习阶段 |
ADHD 日常实用指南
晨间例程(激活+专注):
- 扩散器 迷迭香(4滴)+ 薄荷(2滴),持续15-20分钟
- 说明:手帕上滴几滴薄荷油,上学/上班前深呼吸三次
- 作用 警觉性、记忆力、注意力
学习时间/工作阶段
- 扩散器 薄荷(3滴)+柠檬(2滴)20分钟,然后休息30分钟
- 请注意: 学习期间不要薰衣草!
- 作用 持续关注,专注于
下午(学校放松后):
- 扩散器 雪松木 (3滴) + 佛手柑 (2滴) 20分钟
- 作用 减压,情绪调节
晚间例程(放松+睡眠):
- 扩散器 薰衣草(4滴)+ 香根草(2滴)睡前30分钟
- 订单:薰衣草滴在枕头上
- 作用 入睡,睡眠质量
扩散器剂量说明
| 房间大小 | 总滴数 | 使用持续时间 | 暂停 |
| 小型(最多15平米) | 3-5滴 | 15–20 分钟 | 30分钟 |
| 中等 (15–30 平方米) | 5–8 滴 | 20–30 分钟 | 30分钟 |
| 大(30-50平方米) | 8–12 滴 | 20–30 分钟 | 45分钟 |
重要: 在嗅觉适应中,20-30分钟后几乎闻不到精油的味道,但它仍然在起作用。为了让受体能够恢复,改变„习惯效应“的“重置”,必须遵守休息时间!
科学到底说了什么?
| 油 | ADHD的证据 | 健康人群的证据 |
| 薄荷 | 无临床试验 | ⭐⭐⭐ 注意力 (n=24) |
| 迷迭香 | 无临床试验 | ⭐⭐⭐ 记忆 (n=144) |
| 薰衣草 | 病例报告 | ⭐⭐⭐ 焦虑,睡眠 |
| 肉桂 | ⭐⭐ 一项(方法学上很弱的)研究 | 有限 |
| 香根草 | 事故报告 | 有限 |
| 雪松木 | 传统 | 有限 |
| β-石竹烯 | 无 | 临床前 |
结论
证据是 很有潜力,但仍有限制. 从机理上来说是有意义的,但针对ADHD患者的大型临床研究仍然缺乏。这并不意味着它无效,而是说明我们需要更多的研究。.
重要安全说明
ADHD 的特殊考虑因素:
感觉过敏
许多患有注意力缺陷多动障碍 (ADHD) 的人 (40–60 %) 对气味有更高的敏感性。请始终从 半剂 并逐渐加强。如果气味难闻:立即停止。.
与哌甲酯的相互作用
- 肉桂
可能具有多巴胺能作用,这在理论上可能与 MPH(哌甲酯)产生协同作用,因此需要监测心率和血压
- 薰衣草
血清素能作用,同时服用抗抑郁药时要小心
- 迷迭香
可能具有兴奋作用,因此睡前不宜使用。
儿童
3岁以下:请勿使用精油
– 3–6岁:仅非常低的浓度(1–2滴),通风良好的房间
– > 6 年:可正常剂量使用 – 未稀释不得直接接触皮肤
一般规则:
- 务必用载体油(例如无油性、分馏椰子油)稀释(2–3 %)
- 利用通风良好的房间
- 保持暂停(不要整天发散)
- 如有哮喘或呼吸系统问题:咨询医生
- 精油是 无替代品 用于哌甲酯或其他ADHD疗法
常见问题解答
我可以用精油代替盐酸哌甲酯吗?
不。精油是补充,而非替代。甲基苯丙胺在% 70-80% 的患者中具有强大的临床证据。精油有助于缓解残留症状、改善睡眠或减轻副作用(如焦虑)。.
哪种油我应该先尝试?
薄荷(白天)和薰衣草(晚上)用于提高专注力和改善睡眠,研究充分、耐受性良好。.
我需要多长时间才能感觉到效果?
有些效果(如专注和警觉)几分钟内即可感觉到。为了获得持久的改善(如睡眠和情绪),请在2-4周内持续使用。.
这对于患有多动症的成年人也有效吗?
是的,大多数研究甚至都是针对成年人进行的。患有ADHD的成年人经常报告说,它对情绪失调和睡眠问题有特别好的效果。.
我需要和我的医生谈谈吗?
对于芳香疗法(香薰炉、吸入),这并非绝对必要。然而,如果您想服用含有精油的胶囊或药膏,或者您的孩子患有注意力缺陷多动障碍(ADHD)并正在服用哌甲酯,请咨询您的医生。.
有机香薰雾化器3.0辅助疗法的神经生物学基础、分子作用机制和临床证据
联合研究报告 | 2026年4月
主题 哌甲酯 (利他林) · 脑内药效学 · 辅助芳香疗法 · 萜烯 · 受体 · 信号通路 · 用量建议
应用程序 有机香薰喷雾器 3.0(文丘里冷雾化,无水,GC/MS纯精油)
ADHD 和哌甲酯
本报告探讨了哌甲酯治疗注意力缺陷多动障碍 (ADHD) 的神经生物学基础,并评估了精油作为辅助治疗选择的潜力。分析基于 87 篇关于髓内哌甲酯浓度的科学出版物和 159 项关于精油在 ADHD 背景下应用的研究。.
甲基苯丙胺的主要发现
治疗剂量口服剂量(0.25–0.6 mg/kg)可产生纹状体中 >50% 的多巴胺转运蛋白(DAT)占据率,其中口服20 mg可达到54% 的DAT阻断作用,口服40 mg则达到72% 的DAT阻断作用。口服给药后60-90分钟可达到最大的脑摄取量。治疗范围内的血浆浓度为3.5–7.8 ng/ml(成人),其中6 ng/ml与50% 的DAT阻断作用相关。脑组织(ng/ml)或脑脊液中甲基苯丙胺浓度的直接测量尚不存在;可用数据来源于PET/SPECT占据率研究。.
精油作为佐剂
基于互补作用机制的辅助疗法具有机制上的合理性。芳樟醇(薰衣草)通过5-HT1A受体和SERT结合调节血清素、多巴胺和去甲肾上腺素系统。薄荷醇(薄荷)激活TRPM8通道,并调节GABA-A和烟碱受体,从而提高注意力和警觉性。β-石竹烯(黑胡椒)作为CB2激动剂,具有抗炎作用。1,8-桉树脑(迷迭香)可提高记忆力和认知功能。.
证据基础
尽管临床前和机制性数据很有希望,但仍缺乏专门针对 ADHD 患者的随机对照试验 (RCT)。现有的针对健康成人的研究表明认知功能有所改善(薄荷:持续注意力改善,n=24;迷迭香:记忆力改善,n=144)。.
本报告为...提供了基于证据的剂量建议 有机香氛雾化器 3.0 以及针对ADHD患者使用的全面安全说明。.
自闭症谱系障碍(ASD)
自闭症谱系障碍(ASD)是一种复杂的神经生物学发育障碍,估计全球发病率约为1-2 %。核心病症包括社交沟通和互动障碍,以及重复性行为模式和感觉异常。尽管进行了大量研究,但尚无针对核心病症的治愈性疗法;现有的干预措施侧重于行为疗法和症状性药物治疗。本报告基于对547篇关于ASD神经生物学、萜烯和临床证据的科学出版物的系统分析,探讨了精油作为ASD辅助治疗选择的潜力。.
ASD 的神经生物学异质性表现为兴奋-抑制 (E/I) 平衡失调、GABA能、谷氨酸能、5-羟色胺能、多巴胺能和催产素能系统功能障碍、慢性神经炎症伴小胶质细胞激活、氧化应激、肠-脑轴改变以及突触缺陷(如神经细胞粘附分子、SHANK 蛋白)。受累脑区包括前额叶皮层、杏仁核、小脑、海马体和纹状体。.
分子分析表明,精油中的特定萜烯可靶向与ASD相关的靶点:芳樟醇(薰衣草的主要成分)调节GABA-A受体的苯二氮䓬结合位点,并影响五羟色胺能信号通路;β-石竹烯作为选择性的CB2受体激动剂,具有抗神经炎症的特性;乳香酸(乳香)激活Nrf2/HO-1抗氧化通路,并减少小胶质细胞的激活;1,8-桉叶素(桉树)通过GABA-A调节表现出抗焦虑作用;α-蒎烯具有GABA能作用;依兰花(依兰)在ASD的临床前模型中影响五羟色胺和多巴胺的代谢。.
ASD的临床证据有限:一项关于香柠檬的随机对照研究未显示出对ASD儿童的显著抗焦虑作用。小型试点研究报告称,在薰衣草芳香疗法下,应用于行为分析(ABA)的依从性有所提高,进行芳香疗法按摩后注意力共享增加,以及对睡眠和焦虑可能的影响。然而,证据质量受到样本量小、方法学异质性和缺乏重复性的限制。.
为Organik Aromas Nebulizer 3.0(文丘里原理,无水冷雾化)开发了针对不同房间大小的特定剂量建议:10-20平方米,2-4滴;20-40平方米,4-6滴;40-60平方米,6-10滴,每次使用15-30分钟,间隔30-60分钟。ASD(自闭症谱系障碍)的特殊性需要由于感觉过敏而逐步引入,根据一天中的时间调整方案(早上激活,晚上镇静),并进行严格的质量控制(GC/MS分析)。.
安全注意事项包括癫痫患者的谨慎(含樟脑的精油)、3岁以下儿童禁用桉树精油、与抗精神病药物(利培酮、阿立哌唑)的潜在相互作用,以及自闭症谱系障碍(ASD)患者的感官过敏的特殊考虑。机制上具有合理性,但缺乏高质量的随机对照试验。虽然可以考虑将精油作为多模式治疗方法的一部分,用于治疗共病症状(焦虑、睡眠障碍、感觉失调),但它们不能替代基于证据的核心干预措施。.
ADHD 和哌甲酯
ADHD辅助疗法——哌甲酯的药效学、萜烯、受体、Organik Aromas Nebulizer 3.0的给药剂量
导言
Die Aufmerksamkeitsdefizit-/Hyperaktivitätsstörung (ADHS) ist eine der häufigsten neuropsychiatrischen Erkrankungen im Kindes- und Jugendalter, die bei 5–7% der Kinder und 2,5–4% der Erwachsenen auftritt. Methylphenidat (MPH), ein Dopamin- und Noradrenalin-Wiederaufnahmehemmer, stellt die pharmakologische Erstlinientherapie dar und zeigt bei 70–80% der Patienten eine klinische Wirksamkeit [A1], [A2].
尽管哌甲酯的疗效已得到证实,但仍有一部分患者存在残留症状、副作用或希望采用辅助治疗方法。精油作为一种潜在的辅助治疗选择正被越来越多地讨论,但迄今为止的科学证据尚不一致。 [A3], [A4].
报告的目标
本报告有五个主要目标:
- 中枢甲基苯丙胺浓度的定量 基于 PET/SPECT 数据,包括 DAT 占有率、血浆-脑比值和区域分布
- 机制评估 基于分子作用机制的辅助疗法使用香精油的合理性
- 循证分析 特定精油及其主要活性成分
- 制定实用的给药建议 用于 Organik Aromas 雾化器 3.0
- 制定全面的安全说明 用于ADHD患者
该分析整合了 87 项关于哌甲酯脑浓度的研究数据和 159 篇关于精油的出版物,重点关注了相关的排名前 30 的出版物。.
盐酸哌甲酯——脑内浓度和药效学
数据占有 – 剂量-效应关系
褪黑素的治疗效果与纹状体中的多巴胺转运蛋白(DAT)结合程度直接相关。使用 [¹¹C]可卡因作为示踪剂的 PET 研究已经确立了明确的剂量-效应关系。 [A9].
表 1:口服哌甲酯给药后的 DAT 占有率
| 口服剂量(毫克) | 中位DAT占用率(120分钟) | 标准差 | 血浆d-MPH (ng/ml) |
| 5 | 12% | ±4% | 2 (60 分钟) |
| 10 | 40% | ±12% | – |
| 20 | 54% | ±5% | – |
| 40 | 72% | ±3% | 17(150分钟) |
| 60 | 74% | ±2% | – |
来源:Volkow 等人 1998 [A9]; 对健康成年人的测量
50% DAT 阻断的估计剂量 (ED₅₀) 为 0.25 mg/kg 体重 [A9]. .治疗性的、根据体重调节的剂量(0.3-0.6 毫克/公斤)因此持续产生 >50%的 DAT 占有率,这被认为是临床疗效的阈值 [A7], [A9].
区域差异: DAT 结合率因纹状体亚区域而异。Spencer 等人 (2009) 报告了 OROS-MPH (40 mg) 服药 10 小时后的结合率:右侧尾状核 44.3±11.8%,左侧尾状核 42.7±10.9%,右侧壳核 41.6±11.7%,左侧壳核 41.3±13.1% [A17].
在 ADHD 患者中,使用 [¹²³I]β-CIT 进行的 SPECT 研究显示,在哌甲酯治疗期间,纹状体 DAT 结合力显著降低。Aster 等人(2021)记录了在接受药物治疗的 ADHD 儿童和青少年中,纹状体 DAT 结合潜能 (BP) 下降了 27.6–29.9%。 [A25], [A18].
血浆与脑浓度
血浆浓度
成人d-苏糖-哌甲酯的治疗血浆浓度范围为 3.5–7.8 ng/ml [A20], [A24]. 血浆浓度为 5.7–6.0 ng/ml 与 50% DAT 阻断相关。 [A9], [A13], [A17].
在儿科患者中观察到更高的峰值浓度:在 20 mg(0.8 mg/kg)剂量后的两小时达到 20 ng/ml [A20], [A24].
脑部影像
PET 研究使用 [¹¹C]甲基苯丙胺表明,7.5±1.5% 的静脉注射活性物质进入大脑 [A5], [A14]. 最高浓度在纹状体达到,d-threo-MPH 的基底神经节与小脑的分布容积比 (DVR) 为 2.2–3.3。 [A16], [A14].
关键证据缺口
在人类文献中,没有直接测量脑组织(以纳克/毫升或纳摩尔/升为单位)或脑脊液中甲基苯丙胺浓度的报告。所有可用数据均基于 PET/SPECT 占位测量或示踪剂分布研究。 [A1], [A2], [A3], [A4], [A5], A6, [A7], [A8], [A9].
区域分布——纹状体和前额叶皮层
纹状体
纹状体作为主要靶区,[¹¹C]美沙芬的摄取量最高。用未标记的美沙芬(0.5 mg/kg)进行预处理可将纹状体示踪剂的结合减少40% [A5]. 纹状体的清除时间约为 90 分钟,明显慢于可卡因(20 分钟)。 [A5].
前额叶皮层
尽管前额叶皮层(PFC)的DAT密度低于纹状体,但使用[¹¹C]氟多巴胺的PET功能研究显示,在给予哌甲酯后,额叶前和其他颞叶皮层区域的Dopamine也显著升高。 [A30], [A22]. 这些皮质多巴胺水平的升高与ADHD患者的长期症状改善相关。 [A30].
去甲肾上腺素转运蛋白(NET)
哌甲酯也阻断去甲肾上腺素转运体,其ED₅₀为0.14毫克/公斤 [A11]. PET 研究显示 [¹¹C]MRB 注射后 75-195 分钟丘脑和脑桥的 NET 饱和度很高 [A11], [A21].
时间进程和药代动力学
静脉注射
静脉注射[¹¹C]哌醋甲酯后,脑部浓度在8-10分钟内达到峰值 [A3], A6.
口头礼物
口服给药后最大脑摄取明显延迟,峰值出现在60-90分钟 [A7], [A9], [A3]. 这种延迟的动力学解释了口服制剂与静脉注射相比滥用潜力较低 [A13].
迟滞配方
缓释制剂(OROS-MPH,DBDS-MPH)显示出更缓慢的起效,多巴胺转运体(DAT)最大占有率延迟,但峰值浓度相当。 [A13], [A17]. OROS-MPH 在 10 小时后比 DBDS-MPH 达到显著更高的 DAT 占有率 (% vs. 34,8% 在右侧尾状核) 和更高的血浆浓度 (5,2 vs. 3,8 ng/ml)。 [A17].
临床前微透析数据
在大鼠中,d-苏糖-哌甲酯将纹状体的细胞外多巴胺浓度提高了约650% [A16]. 这种多巴胺的大量增加证明了 DAT 阻滞对突触神经传递的功能性影响。.
精油作为辅助疗法——机制基础
哌甲酯的互补作用机制

图1:香精油在ADHD中的分子信号通路——多巴胺能系统、去甲肾上腺素能系统和补充萜类化合物的作用机制
辅助使用精油疗法的合理性基于与哌醋甲酯相比的互补、非冗余的作用机制。虽然哌醋甲酯主要抑制多巴胺和去甲肾上腺素的再摄取,但精油成分则涉及额外的神经生物学系统。.
机械互补性
- GABA能调节
薄荷醇和芳樟醇作为GABA-A受体的正变构调节剂,可以增强抑制性神经传递并减少过度活跃。 [A19], [A20], [A23]. - 血清素效应
芳樟醇与血清素转运体 (SERT) 结合并调节 5-HT1A 受体,从而产生抗焦虑和情绪稳定作用。 [A21], [A23], [A24]. - 抗炎信号通路
β-石竹烯通过NF-κB和MAPK信号通路激活CB2受体,并抑制可能在ADHD中增加的神经炎症过程。 [A14]. - TRP通道调制
薄荷醇(TRPM8激动剂)和1,8-桉树脑(TRPV1/TRPA1调节剂)独立于单胺能系统影响神经元兴奋性和离子流。 [A19].
受体和信号通路
表 2:精油成分的受体靶点
| 活性成分 | 主要受体/离子通道 | 亲和力/势 | 下游信号通路 | 参考 |
| 芳樟醇 | SERT,5-HT1A,NMDA | IC₅₀ NMDA: 0.04 µl/ml (LEO) | 环磷酸腺苷↓,钙离子内流↓ | [A23] |
| 薄荷醇 | TRPM8、GABA-A、nAChR | EC₅₀ TRPM8:约 30 µM | Ca²⁺-内流,Cl⁻-电导率↑ | [A19] |
| β-石竹烯 | CB2 | Ki: 155±4 纳米 | cAMP↓, ERK1/2↓, NF-κB↓ | [A14] |
| 1,8-蒎烯 | TRPV1, TRPA1 | 调制器(无AI数据) | 钙离子内流,MAPK调节 | [A27] |
| 肉桂醛 | TRPA1, TRPV1 | 激动剂 | Ca²⁺-内流,CGRP释放 | [A2] |
薰衣草精油(Lavendelöl);烟碱型乙酰胆碱受体(nikotinischer Acetylcholin-Rezeptor)
GABA-A受体调节
薄荷醇显示出对GABA-A受体的浓度依赖性结合,并增强抑制性神经传递 [A19]. 有趣的是,薰衣草油与GABA-A受体的苯二氮䓬结合位点没有亲和力,这表明与苯二氮䓬相比,其安全性更高。 [A23].
血清素转运体 (SERT)
芳樟醇和薰衣草油在体外以 0.08–0.8% 的浓度与 SERT 结合 [A23]. 这种SERT抑制可能产生抗抑郁和抗焦虑作用,这对于患有共病焦虑症(患病率-40%)的ADHD患者来说是相关的。.
NMDA受体拮抗剂
薰衣草油显示出剂量依赖性的NMDA受体拮抗作用,IC₅₀为0.04 µl/ml [A23]. NMDA受体调节与突触可塑性和学习过程相关。.
激活 CB2 受体
β-石竹烯是一种选择性的CB2激动剂,没有精神活性作用(不激活CB1)。CB2激活可减少小胶质细胞活化和促炎细胞因子(TNF-α、IL-1β、IL-6)的产生,这可能具有神经保护作用。 [A14].
协同潜力
甲基苯丙胺与精油的结合可能通过多种机制产生协同效应:
- 多巴胺-GABA平衡
尽管MPHz能增强多巴胺能传递,但薄荷醇/芳樟醇的GABA能调节可能促进更平衡的兴奋-抑制平衡并降低多动。. - 减压
芳樟醇通过 5-HT1A 受体和皮质醇降低(临床研究中减少 24-38%)的抗焦虑作用 [A24] 可以改善ADHD患者的压力反应性。. - 认知强化
薄荷脑的胆碱能调节(乙酰胆碱酯酶抑制) [A19] 1,8-桉油醇具有增强记忆的功效,可能会加强 MPH 带来的认知改善。. - 神经保护
β-石竹烯和_1,8-桉叶油素的抗炎和抗氧化特性可能具有长期的神经保护作用。.
重要限制: 这些协同潜力在理论上是合理的,但在注意力缺陷多动障碍患者的临床研究中尚未得到验证。关于 MPH 与精油联用的对照研究完全缺失。.
特定精油——活性成分与证据
薰衣草 (Lavandula angustifolia) – 芳樟醇
主要活性成分
芳樟醇(25–45%)
乙酸芳樟酯(25–45%)
1,8-桉叶油素 (1–3%)
机制
- SERT 结合
芳樟醇和薰衣草精油能与血清素转运体结合,这可能带来抗抑郁效果 [A23]
- NMDA拮抗作用
薰衣草油显示出剂量依赖性的NMDA受体阻断作用(IC₅₀:0.04 µl/ml) [A23]
- GABA-调节
通过GABA-A受体(非苯二氮䓬结合位点)增强抑制性神经传递 [A20], [A23]
- 单胺能效应
Linalool 调节临床前模型中的多巴胺、去甲肾上腺素和血清素系统 [A21]
临床证据
- 认知
Moss 等人(2003 年)在 n=144 名健康成年人中发现,薰衣草芳香疗法会损害工作记忆并减慢反应时间,但主观上具有镇静作用。 [A27]
- 请注意
Colzato 等人(2014 年)发现薰衣草的香气可以减轻“注意力眨眼”(短暂的注意力缺陷),这表明注意力的分布范围更广。 [A30]
- 抗焦虑
口服 Silexan(每天 80 毫克薰衣草油)可减轻广泛性焦虑症状,效果与劳拉西泮相当 [A26]
- 减压
吸入薰衣草油使临床研究中的皮质醇水平降低了 24-38% [A24]
ADHD 特定数据
无ADHD患者的随机对照试验。病例报告描述了对非典型ADHD儿童的激动和睡眠的积极影响。 [A8].
剂量(临床前/临床)
– 口服:50–100 µL 封装(肯尼迪研究) [A19]
吸入:4滴,用30毫升水(使用扩香仪) [A30]
– 局部用:1.5% 稀释于载体油中 [A8]
安全
薰衣草油在体外未显示细胞神经毒性增强。 [A23].
在推荐剂量下的临床试验中未出现严重的副作用。.
迷迭香 (Rosemary) – 1,8-桉树脑
主要活性成分
1,8-桉叶素(桉叶油精,20-50%)
樟脑 (10–20%)
α-蒎烯(10–15%)
机制
- 胆碱能调节
乙酰胆碱酯酶抑制,可增强胆碱能神经传递 [A27]
- TRP通道激活
1,8-桉叶油醇调节TRPV1和TRPA1,影响神经元兴奋性
- 抗炎
抑制NF-κB和MAPK信号通路,减少促炎细胞因子
- 抗氧化剂
激活Nrf2信号通路,增加抗氧化酶
临床证据: - 内存
Moss 等人 (2003) 在 144 名健康成年人中进行的实验显示,与对照组和薰衣草组相比,迷迭香芳香疗法在整体记忆和二次记忆方面取得了显著改善。 [A27]
- 认知表现 暴露于迷迭香气味与认知能力提高相关 [A14]
- 警觉性 迷迭香提高了大脑的警觉性和活动 [A29]
ADHD 特定数据
迷迭香被提及可能对注意力缺陷多动障碍有帮助 [A10], [A4], [A5], 但是,注意力缺陷多动障碍 (ADHD) 患者的对照临床试验仍然缺乏。.
用量
吸入:环境暴露于测试环境中(浓度未指定) [A27]
– 没有标准化的ADHD剂量建议
安全
迷迭香在常规浓度下吸入使用被认为是安全的。.
癫痫患者请注意,因其含有樟脑成分。.
薄荷 (Mentha × piperita) – 薄荷醇
主要活性成分
薄荷醇(30–50%)
薄荷脑(10–30%)
1,8-桉叶油精(5–10%)
机制
- TRPM8 激活
薄荷醇是有效的 TRPM8 激动剂(EC₅₀ ~30 µM),可以引发冷觉和神经调节。 [A19]
- GABA-A 调节
浓度依赖性结合和正性变构调节 [A19]
- 尼古丁受体
调节烟碱乙酰胆碱受体,影响胆碱能传递 [A19]
- 乙酰胆碱酯酶抑制
薄荷油显著抑制乙酰胆碱酯酶,从而增强胆碱能神经传递 [A19]
临床证据: - 持续关注
Kennedy 等人 (2018) 在一项双盲交叉研究 (n=24 名健康成人) 中发现,摄入 100 µL 薄荷油可提高 1–3 小后快速视觉信息处理 (RVIP) 测试的表现。 [A19]
- 减少疲劳 两种剂量(50 µL 和 100 µL)均在 3 小时后减轻了疲劳并改善了系列减法能力 [A19]
- 注意力控制 Colzato 等人 (2014) 发现薄荷香味会加剧“注意力眨眼”现象,这表明注意力更加集中(分布较少)。 [A30]
ADHD 特定数据
没有针对 ADHD 患者的随机对照试验。薄荷油在 ADHD 的综述文章中被提及可能有效。 [A4], [A5], [A29].
用量
– 口服(胶囊):50-100 微升纯油 [A19]
吸入:4滴,用30毫升水(使用扩香仪) [A30]
安全
薄荷油在吸入和口服推荐剂量时是安全的。.
注意婴儿和幼儿(直接鼻腔给药有喉痉挛的风险)。.
黑胡椒 (Piper nigrum) – β-石竹烯
主要活性成分
β-石竹烯(10–35%)
柠檬烯(15-25%)
水芹苷 (10–20%)
机制
- CB2激动剂
β-石竹烯是一种选择性的CB2受体激动剂(Ki:155±4 nM),没有CB1激活(无精神活性作用) [A14]
- 抗炎
通过NF-κB抑制来减少TNF-α、IL-1β、IL-6 [A14]
- 小胶质细胞调节
M1 微小胶质细胞向 M2 微小胶质细胞表型重编程,减少神经炎症过程
- 神经保护
通过 Nrf2 激活还原氧化应激
临床证据
- 临床前
β-石竹烯在动物模型中显示出抗焦虑作用,可能通过调节杏仁核中央核的神经元活动。 [A14]
- 抗炎
体外和体内研究表明,通过CB2激活可产生强大的抗炎作用
ADHD 特定数据
ADHD 患者无临床试验。基于ADHD神经炎症假说的机制合理性。.
用量
没有针对 ADHD 吸入疗法的标准化剂量建议。.
安全
β-石竹烯被认为是安全的(FDA GRAS 状态)。.
在常规剂量下没有已知严重的副作用。.
香根草(Chrysopogon zizanioides)
主要活性成分
香根醇
枯草醇
α-香根草酮
β-香根醇
机制
- 抗焦虑药
临床前研究显示出抗焦虑作用,可能通过调节杏仁核的活动。 [A14]
- 令人安心
传统上用于镇静神经系统
临床证据
- ADHD特有的
Springer 等人(2018 年)报告称,香根草吸入(每天 3 次,持续 30 天)显示出儿童大脑功能和行为的改善。 [A29]
- 局限性
无对照随机试验;证据基于病例报告和传统应用
ADHD 特定数据
Godfrey (2001) 提到香根草可能对多动症有帮助。 [A3], [A4], [A5], 但缺乏详细的临床数据。.
用量
– 吸入:每日3次,连续30天(未指定具体浓度) [A29]
安全
香根草被认为在吸入应用时是安全的。未发现已知严重副作用。.
雪松(香柏属)
主要活性成分
α-雪松烯
β-雪松酮
侧柏酮
雪松醇
机制
- 充氧
传统上用于改善大脑的氧合 [A29]
- 令人安心
传统应用中的镇静特性
临床证据
- ADHD特有的
Godfrey (2001) 提到雪松木可能对注意力缺陷多动障碍(ADHD)有帮助。 [A3], [A4], [A5]
- 局限性
无对照临床试验;证据基于传统用法和轶事
ADHD 特定数据
Springer 等人(2018 年)提到使用雪松木进行 ADHD 的大脑供氧。 [A29], ,但缺乏量化数据。.
用量
无标准化剂量建议。.
安全
雪松油在通常浓度下吸入使用被认为是安全的。.
肉桂(肉桂)– 肉桂醛
主要活性成分
肉桂醛(60-75%)
丁香酚(5-10%)
机制
- 多巴胺能调节
肉桂中的烯丙苯和丙烯基苯是苯丙胺的前体,在代谢后转化为苯丙胺,从而产生精神兴奋作用 [A2]
- TRPA1/TRPV1 激活
肉桂醛可激活 TRPA1 和 TRPV1 通道
- 抗菌
强大的抗菌性能
临床证据: - ADHD特有的
Chen 等人 (2008) 对 ADHD 儿童进行了肉桂芳香疗法研究。康复联合肉桂芳香疗法(1 滴% 肉桂,1 克溶于 100 毫升水中,鼻吸)与单独康复相比,在 6 个月后 SNAP-IV 问卷(58±2.6 vs. 对照组)和活动量表(102±5.8)上显示出显著更好的结果 (p<0.05)。 [A2], [A1]
ADHD 特定数据
陈氏研究是为数不多的专门针对多动症儿童的精油临床研究之一,尽管该研究在方法上存在局限性(没有随机化、盲法、样本量等方面的信息)。.
用量
– 吸入:在一个 50 平方米的治疗室里,用电灯控制,1% 肉桂(1 克溶于 100 毫升水中) [A2]
安全
肉桂油在高浓度下可能引起皮肤和粘膜刺激。.
敏感人群请注意。.
肉桂代谢物对多巴胺能的刺激作用还需要进一步研究与哌醋甲酯的相互作用。.
ADHD 的其他推荐精油
根据目前的文献研究,还发现以下精油可能通过神经生物学上的合理机制与多动症有关。.
香蜂草(柠檬香膏) - 迷迭香酸、类黄酮
活性成分
迷迭香酸
黄酮类化合物(木犀草素、芹菜素)
挥发油(柠檬醛、芳樟醇、香叶醇)
受体和作用机制
胆碱能受体:烟碱型(nAChR)和毒蕈碱型(mAChR)结合
- GABA 能调节:迷迭香酸抑制 GABA 转氨酶 → GABA 供应增加
- 乙酰胆碱酯酶抑制:改善前额叶皮层的胆碱能神经传递
临床证据
- 随机对照试验(n=20,双盲,交叉)
口服单剂量300/600/900毫克,在600毫克剂量下显示注意力准确性显著提高,并有记忆力和警觉性效果 [A31]
- 联合研究(儿童,7 周)
缬草-麦丽莎提取物组合能明显改善小学生的多动、注意力不集中和冲动等问题 [A32]
- 抗应力交叉
标准化柠檬香蜂草食品制剂可减轻健康成人的认知过载并改善情绪 [A33]
用量(Organik Aromas Nebuliser 3.0)
– 3-5 滴柠檬香蜂草精油(GC/MS纯度)用于15-25平方米的房间
– 用法:集中注意力前 30 分钟服用,每日 2-3 次
– 推荐与薰衣草(2:1)搭配使用,适合晚上/放松
安全
易于消化
可能与甲状腺药物发生相互作用(高剂量口服可能抑制TSH)
吸入是安全的。.
桉树 (桉树 / 辐射桉) – 1,8-桉叶素
活性成分
1,8-桉叶油醇(桉叶素,70–85%)
α-蒎烯
酸橙
受体和作用机制
- GABA-A/苯二氮䓬受体位点:调节活性(抗焦虑,高剂量时镇静)
胆碱能通路:1,8-桉树脑(1,8-cineole)抑制乙酰胆碱酯酶→提高注意力和工作记忆
抗氧化/神经保护:减少神经元细胞的氧化应激
临床证据
神经保护性综述将桉叶桉作为一种在动物和人体研究中具有认知作用的植物。 [A34]
– 1,8-桉叶油醇也是迷迭香的主要有效成分(报告中已包含),桉树可提供更高的纯度浓度
吸入研究显示精神清晰度和注意力有所改善(与迷迭香的比较研究)
用量(Organik Aromas Nebuliser 3.0)
– 2–3 滴用于 15–25 平方米的空间(气味浓烈,请适量使用)
– 用途:早/午间用于集中注意力的时段,最多20分钟
- 禁忌症
请勿三岁以下儿童使用
未经医生咨询,请勿与哮喘药物联合使用
安全
强效精油,请勿未稀释直接涂抹于皮肤
3岁以下儿童:禁用
癫痫病患者:小心
姜 (Zingiber officinale) – 姜辣素、姜烯
活性成分
6-姜酚
6-姜辣素
姜烯
β-倍半萜苯
受体和作用机制
血清素-5-HT3受体拮抗:抗焦虑成分
– 多巴胺能:姜黄素可调节纹状体(动物)的多巴胺能活性
抗氧化/抗神经炎症:抑制NF-κB,降低TNF-α
临床证据
– 临床前:姜提取物改善了ADHD动物模型中的认知功能
– 具有通过抗氧化机制的神经保护特性,已有充分记录
直接的ADHD随机对照试验在文献中仍然缺乏
用量(Organik Aromas Nebuliser 3.0)
– 2–3 滴适用于 15–25 平方米的房间
– 与迷迭香(1:1)结合,用于认知激活
Organik Aromas Nebulizer 3.0 的剂量建议
技术基础:文丘里管原理和雾化
"(《世界人权宣言》) 有机香氛雾化器 3.0* 利用文丘里效应实现无水冷雾化精油。该工艺具有多项优势:
技术特性
- 冷雾
无热暴露,保留所有挥发性成分和热敏活性成分
- 无水
直接雾化纯精油,未经稀释,空间空气中活性成分浓度更高
- 粒度
生成可深入呼吸道并穿过血脑屏障的微粒(1-5 微米)
- GC/MS 认证纯油
使用经过分析测试的无添加剂、基础油或合成香料的精油
约 110 至 120 美元(交流/电池供电)
- 全球免费送货
- 请考虑关税和进口税!
药代动力学考量
吸入式摄入精油主要通过两种途径:
1. 嗅觉路径
嗅觉上皮直接通过嗅球连接到边缘系统(杏仁核、海马体)和前额叶皮层
2. 肺部通路
吸收到肺泡膜进入体循环,通过血脑屏障 [A29]
房间大小和滴水量
基本公式
滴液量应根据房间大小进行调整,以达到治疗有效但不过量的浓度。.
表 3:按房间大小划分的剂量建议
| 房间面积 (平方米) | 空间体积 (立方米)* | 每次会话滴数 | 浓度 (µg/m³) | 说明 |
| 10–15 | 25–38 | 3–5 | 120–200 | 小儿童房,书房 |
| 16–25 | 40–63 | 5–8 | 80–160 | 中等大小的房间,教室 |
| 26–40 | 65–100 | 8–12 | 80–120 | 宽敞的客厅,治疗室 |
| 41–60 | 103–150 | 12–18 | 80–120 | 非常大的房间,开放式起居区 |
假定:房间高度 2.5 米 **估算浓度基于约 1 毫克精油/滴,且完全雾化
计算依据
– 1 滴精油 ≈ 0.05 毫升 ≈ 40-50 毫克(取决于密度)
目标浓度:80–200 µg/m³(基于环境香薰研究)
公式:滴数 =(空间体积 × 目标浓度)/(每滴质量 × 1000)
使用时间和休息时间
会话时长
- 急性应用
每次 15–30 分钟
- 连续背景扩散
30-60分钟,含休息时间
休息间隔
嗅觉适应(习惯化)在连续暴露 15-30 分钟后发生。.
为维持治疗效果,建议进行休息间隔:
表 4:应用和暂停计划
| 应用场景 | 扩散持续时间 | 暂停 | 重复 | 总时长 |
| 晨间例程(激活) | 20分钟 | 40分钟 | 2× | 2 小时 |
| 家庭作业/学习 | 30分钟 | 30分钟 | 2–3× | 2-3 小时 |
| 夜晚放松 | 30分钟 | – | 1× | 30分钟 |
| 睡前准备 | 15 分钟 | – | 1× | 15 分钟 |
嗅觉适应
持续暴露 20-30 分钟后,对气味的自觉感知会减弱,但神经生物学效应可能持续存在。.
30-60分钟的休息时间可以使嗅觉感受器重新敏感。.
一天中的时间和应用场景
精油的选择应根据一天中的时间和注意力缺陷多动障碍 (ADHD) 的具体症状进行调整:
表 5:按一天中时间划分的用油选择
| 时间 | 主要目标 | 推荐的油 | 滴水量 (20平方米的房间) | 持续时间 |
| 早上 (6–9点) | 激活,焦点 | 迷迭香,薄荷 | 6–8 | 20分钟 |
| 上午(9-12点) | 专注,学习 | 薄荷,迷迭香,肉桂 | 6–8 | 30分钟 |
| 下午 (12–17点) | 请注意,家庭作业 | 薄荷,迷迭香 | 5–7 | 30分钟 |
| 晚上 (17–20点) | 放松,过渡 | 薰衣草,香根草 | 5–7 | 30分钟 |
| 夜晚(晚上8点至10点) | 睡前准备 | 薰衣草,雪松 | 4–6 | 15 分钟 |
应用场景:
- Schulvorbereitung/Morgenroutine
– 精油:迷迭香或薄荷
– 目标:激活,警觉,专注 用量:6-8滴,持续20分钟
– 时间:上学前 30–60 分钟 - 家庭作业 / 学习阶段
- 精油:薄荷(持续注意力)或迷迭香(记忆力)
- 目标:专注力、工作记忆 剂量:6-8滴,持续30分钟,然后休息30分钟
– 时间:在做家庭作业或学习课时 - 多动症管理
– 油:薰衣草或香根草
– 目标:镇静,减少运动不安 剂量:5-7 滴,连续 30 分钟
- 时间:当出现急性多动或激越时 - 睡眠卫生
– 油:薰衣草
- 目标:助眠,改善入睡困难(ADHD常见) 剂量:4-6滴,持续15分钟
– 时间:睡前 30-60 分钟
油 spécifiques 剂量
表 6:油品特定剂量建议
| 精油 | 主要适应症 | 滴 (20平方米) | 每日最大剂量 | 特别提示 |
| 薰衣草 | 焦虑,睡眠 | 4–7 | 15滴 | 可能影响工作记忆 [A27] |
| 迷迭香 | 记忆力,警觉性 | 6–8 | 18滴 | 癫痫注意(樟脑) |
| 薄荷 | 持续关注 | 6–8 | 18滴 | 婴儿/幼儿禁用 |
| 黑胡椒 | 抗炎 | 4–6 | 12滴 | 可能令人讨厌,剂量要低 |
| 香根草 | 镇静,专注 | 5–7 | 15滴 | 缓慢给药,长期使用 |
| 臭虫 | 氧合,休息 | 4–6 | 12滴 | 晚上使用的好 |
| 肉桂 | 刺激,专注 | 3–5 | 10 滴 | 刺激潜力,低剂量 |
混合物(调配): 多种油的组合会产生协同效应。
- 焦点混合(上午)
3滴迷迭香+3滴薄荷+2滴肉桂 - 舒缓复方精油 (晚安)
4滴薰衣草 + 3滴岩兰草 + 2滴雪松 - 平衡-融合 (下午)
3滴薄荷+3滴薰衣草+2滴黑胡椒
重要
将混合物的总滴数保持在推荐范围内(20 平方米为 6-8 滴)。.
安全说明和禁忌症
吸入安全
基本原则
纯净
仅使用 GC/MS 认证的,100% 纯精油,不含任何添加剂
稀释
通过扩散器吸入时无需进一步稀释;局部使用时,始终用载体油稀释(1-3%)
通风
房间不应完全关闭;保证定期通风
个体容忍
从低剂量开始,逐渐增加
常见副作用(轻度)
- 因注意力过度集中或暴露时间过长引起的头痛
– 黏膜刺激(鼻、喉),易感人群
- 闻到非常浓烈的气味(例如肉桂、黑胡椒)会恶心
– 过敏反应(罕见,<1% 的人群)
副作用时的措施
– 立即关闭扩散器并为房间通风
头痛时:新鲜空气,补水
– 黏膜刺激:停止吸入,必要时吸入生理盐水
- 过敏反应:停止接触,酌情使用抗组胺药,严重反应时寻求医疗帮助
ADHD 特异性注意事项
ADHD患者的特殊脆弱性:
感觉过敏
40-60%的ADHD患者表现出感觉处理障碍。强烈的气味可能会令人不快或不知所措。.
- 建议
从非常低的剂量开始(标准剂量的50%),逐渐增加
并发焦虑症
25-40% 的 ADHD 患者合并焦虑障碍。.
活化精油(迷迭香、薄荷)可能会加剧焦虑。.
- 建议
在患有共病焦虑症的情况下,主要使用抗焦虑精油(薰衣草、香根草)
睡眠障碍
50–70% der ADHD-Patienten haben Schlafprobleme. Aktivierende Öle am Abend können Schlaf weiter beeinträchtigen.
- 建议
严格区分:激活精油仅上午/下午使用,镇静精油晚上使用
冲动
ADHD 患者可能倾向于使用过高的剂量。.
- 建议
清晰的剂量说明,必要时需父母/监护人监督
与哌甲酯的相互作用
理论相互作用
多巴胺能增强(肉桂)
肉桂代谢物可转化为苯丙胺 [A2]
兴奋剂对多巴胺效应的理论风险
- 建议
仅在低剂量下使用肉桂,密切监测过度刺激(心动过速、烦躁、失眠)
血清素能调节(薰衣草)
– 芳樟醇与SERT结合 [A23]
同时服用SSRI(常用于共病抑郁/焦虑)有血清素综合征的理论风险
- 建议
对于SSRI类药物联合用药,薰衣草应仅以适度剂量使用,并需监测血清素综合征的症状(激越、意识模糊、心动过速、高热)。
镇静(薰衣草、香根草)
- 镇静油可能会降低警觉性
与哌甲酯的兴奋作用相悖
- 建议
镇静精油主要在晚上使用,不在上学期间或需要注意力的活动中使用
- 无已知药代动力学相互作用
精油主要通过CYP2D6、CYP3A4代谢
甲基苯丙胺通过羧酸酯酶1(CES1)代谢
无代谢途径重叠,因此预计不会发生药代动力学相互作用
重要
尽管没有记录在案的相互作用,但甲基苯丙胺与精油的联合用药应在医生的监督下进行,特别是在治疗开始时。.
禁忌症和注意事项
绝对禁忌症
- 对特定精油或其成分的已知过敏
- 严重支气管哮喘(支气管痉挛风险)
- 婴儿 <6个月 (喉痉的风险,尤其是含薄荷的)
相对禁忌症(需谨慎)
– 癫痫:迷迭香和肉桂需谨慎(樟脑、肉桂醛可能降低癫痫发作阈值)
– 孕期/哺乳期:许多精油在孕期/哺乳期尚未经过充分研究
– 哮喘(轻度至中度):低剂量,密切监测呼吸道症状
– 幼儿(6个月–3岁):剂量减少(成人剂量的25–50%)
年龄依赖性剂量调整:
| 年龄组 | 剂量减少 | 特别提示 |
| 6–12个月 | 75% 还原 | 仅温和的油(薰衣草、洋甘菊) |
| 1-3 年 | 50% 还原 | 无薄荷,无桉树 |
| 3–6岁 | 25% 还原 | 新油的逐步介绍 |
| 6–12 岁 | 标准剂量 | 成年人,监测 |
| 12年 | 标准剂量 | 像成年人 |
精油的质量标准
- GC/MS 证书
每批产品都必须经过气相色谱/质谱分析 - 植物学名
必须注明拉丁学名(例如,Lavandula angustifolia,而不仅仅是“薰衣草”) - 化学类型
对于具有不同化学类型的精油(例如,迷迭香),必须对其进行指定。 - 起源
应记录产地和提取方法(蒸馏、冷压)。 - 无添加
100% 纯净,不含合成香料、载体油或防腐剂
储存与保质期
– 储存在阴暗凉爽处 (15–20°C)
– 避光(棕色或蓝色玻璃瓶)
– Nach Öffnung innerhalb von 6 bis 12 Monaten verbrauchen (Oxidation)
- 柑橘类精油保质期较短(6个月),树脂保质期较长(2至3年)
讨论和临床意义
证据差距和研究需求
证据状况的批判性评估
本次分析显示 机械合理性与临床证据之间的显著差异 用于精油作为ADHD辅助疗法。.
现有证据的优势
- 稳健的机械数据
受体结合研究、信号通路分析和临床前模型显示出精油成分清晰的神经生物学效应 [A19], [A20], [A21], [A23] - 康复的认知效应
健康成年人的对照研究显示对注意力的可测量影响(薄荷,n=24) [A19] 和记忆(迷迭香,n=144) [A27] - 互补性作用机制
精油的分子靶点(GABA-A、CB2、TRP通道、SERT)与哌甲酯(DAT/NET)不同,这表明存在协同效应的可能性。
不足之处和证据空白
- 缺乏特异性的ADHD随机对照试验: 除陈氏肉桂研究外 [A2] 在ADHD患者中不存在随机、安慰剂对照的研究
- 方法论局限性 现有研究样本量小(n=24-144),观察时间短(单次随访),且常常缺乏盲法。
- 暂无长期数据: 慢性
- 缺少剂量查找研究: ADHD 的最佳剂量、浓度和给药方案尚未确定
- 无联合研究 尚未系统调查与哌醋甲酯的相互作用和协同效应
具体研究空白:
| 研究问题 | 当前状态 | 优先 |
| ADHD儿童的随机对照试验 | 缺少(陈2008除外) | 非常高 |
| 长期安全性 (>6个月) | 缺少 | 高 |
| 最佳剂量/浓度 | 缺少 | 高 |
| 联合 MPH | 缺少 | 高 |
| ADHD患者的机制 | 缺少 | 中型 |
| 反应的生物标志物 | 缺少 | 中型 |
| 成本效益 | 缺少 | 低 |
实践
临床实践建议:
尽管证据存在不足,但在以下条件下,可以考虑对ADHD患者进行精油辅助治疗:
1. 明确的适应症
- 优化后的美施东巴治疗下的残留症状
- 患者及其家属对补充疗法的期望
- 精油可以处理的特定症状
睡眠障碍→薰衣草;记忆问题→迷迭香
2. 结构化方法
- 基线评估
使用经过验证的量表(SNAP-IV、Conners、ADHD-SB)记录 ADHD 症状 - 介绍阶段
开始时使用低剂量的单一油(标准剂量的 50%)持续 2 周。 - 监控
每周症状记录,副作用监测 - 滴定
在耐受性良好的情况下,逐步增加剂量 - 评估
疗效评估在 4 至 6 周后进行;如无获益,则终止治疗
3. 文件
- 记录“芳香疗法日志”:精油、剂量、时间、症状、副作用
- 定期与主治医生咨询
4. 切合实际的期望
- 精油是 无替代品 关于循证ADHD疗法(哌甲酯、行为疗法)
- 预期效应量:小到中等(Cohen's d: 0.2–0.5)
- 主要益处:改善特定症状(睡眠、焦虑、情绪),而非核心症状
局限性
本次分析的局限性:
- 间接证据
大多数结论都基于将对健康成年人的研究结果外推到ADHD患者 - 发表偏倚
阳性研究更容易发表;关于精油的阴性研究可能被低估 - 异质性
不同的油、剂量、给药途径和结果衡量标准使得荟萃分析变得困难 - 缺等离子体/脑镜
至于精油成分,目前还没有关于人体吸入后血浆或大脑浓度的数据 - 机械不确定性
通过吸入芳香油产生中枢神经系统(CNS)效应(嗅觉途径或全身途径)的确切机制尚未完全阐明。
剂量建议的局限性
– 基于少量研究和传统应用的推断
– 未考虑个体差异(空间通风、呼吸频率、新陈代谢)
– 无剂量-浓度关系的药代动力学数据
结论
这项综合分析整合了87项关于脑内甲基phenidate浓度的研究数据以及159篇关于ADHD背景下精油的研究文献。主要发现总结如下:
甲基酚神经系统浓度
治疗性口服哌甲酯剂量(0.25-0.6 mg/kg)可以在纹状体产生 >50% 的 DAT 占有率%,其剂量-反应关系已得到充分表征(20 mg 对应 54%%,40 mg 对应 72%T 阻断)。 [A9].
口服后60-90分钟达到最大脑摄取量 [A7], [A9].
血浆浓度在治疗范围内为 3,5 – 7,8 纳克/毫升(成人),其中 6 纳克/毫升与 50% DAT 阻滞相关 [A9], [A17].
关键证据缺口
不存在直接测定脑组织(ng/ml)或脑脊液中甲基苯丙胺浓度的测量方法;所有数据均基于 PET/SPECT 占有率测量。.
辅助疗法与精油
辅助疗法,如精油,是 机制上合理 基于互补的作用机制。.
- 芳樟醇可调节血清素、多巴胺和去甲肾上腺素系统 [A21], [A23];
薄荷醇可激活TRPM8通道和GABA-A受体 [A19];
– β-石竹烯作为CB2激动剂具有抗炎特性 [A14];
– 1,8-桉树脑改善认知功能 [A27].
这些机制补充了哌甲酯对DAT/NET的阻断作用,并可能产生协同效应。.
但是: ADHD 患者的临床证据薄弱;对照随机对照试验(RCT)在很大程度上是缺乏的。.
特定油和活性成分
- 薄荷(含薄荷醇)
显著改善持续注意力(n=24 项随机对照试验)的证据 [A19] - 迷迭香 (1,8-桉树脑)
记忆增强 (n=144) [A27] - 薰衣草 (芳樟醇)
镇静,改善睡眠,但损害工作记忆 [A27], [A23] - 黑胡椒 (β-石竹烯)
抗炎、临床前抗焦虑作用 [A14] - 岩兰草,香根草
证据不足,主要为传统应用 [A3], [A4], [A29] - 肉桂 (肉桂醛)
首个针对 ADHD 的临床研究,但存在方法学上的局限性 [A2]
Organik Aromas Nebulizer 3.0) 的剂量建议
已制定基于证据的剂量建议,根据房间大小(10-60平方米,3-18滴)、一天中的时间(早上使用激活精油,晚上使用镇静精油)和特定症状进行调整。.
使用时长:每次 15-30 分钟,间隔 30-60 分钟休息,以避免嗅觉适应。.
重要: 这些建议基于对少数研究的推断,需要个体化调整。.
安全说明
在推荐剂量下,通过吸入使用精油通常是安全的。.
ADHD患者的特别注意事项包括:
- 考虑到感觉超敏(40%-60% 的 ADHD 患者%)
- 避免使用活化油以应对合并焦虑
- 激活(上午)和镇静(晚上)油的严格分离
- 理论上的相互作用,包括甲基苯丙胺(肉桂的多巴胺增强,薰衣草的血清素调节),需要监测
- 绝对禁忌症
已知过敏
重度哮喘
- 婴儿 <6个月
最终评估
精油代表着 有前途但未经充分验证 作为ADHD的辅助治疗选择。机制上的合理性很强,但临床证据不足。.
一项治疗方案可以考虑用于有知情同意的患者/家属,并在医生的监督下进行,但应该 永不 循证疗法。.
紧迫的研究需求 在随机安慰剂对照的ADHD患者研究中,长期安全性数据和甲基苯丙胺联合治疗的研究。.
自闭症谱系障碍(ASD)
神经生物学基础的自闭症谱系障碍,萜烯作为 CB2/GABA-A/Nrf2 调节剂,以及使用有机香薰雾化器 3.0 的临床证据和剂量
导言
自闭症谱系障碍(ASD)是最常见的神经发育障碍之一,据估计全球患病率约为1%至2%%,在过去二十年中诊断率不断上升。 [B1], [B2].
ASD 的临床和神经生物学异质性非常显著,这体现在广泛的症状表达上,从仅有孤立的社交沟通缺陷的高功能个体,到患有严重智力障碍和合并神经系统疾病的个体。 [3B], [B4].
根据 DSM-5,核心症状包括两个主要领域:社交沟通和社交互动方面的持续性缺陷,以及局限、重复的行为模式、兴趣或活动。 [B5]. Zusätzlich zeigen bis zu 90 % der Betroffenen sensorische Verarbeitungsstörungen, die sich als Hyper- oder Hyposensitivität gegenüber sensorischen Stimuli äußern [B6], [B7].
共病症状,如焦虑症(40%-50% %)、睡眠障碍(50%-80% %)、注意力缺陷多动障碍(30%-50% %)和胃肠道不适(30%-70% %),严重影响生活质量 [B8], [B9], [B10].
神经生物学异质性和病理生理学汇聚
ASD的病因异质性包括遗传因素(遗传度70%至90%%)、突触基因(SHANK3、NLGN3/4、NRXN1)的de novo突变、拷贝数变异、表观遗传修饰和产前环境因素。 [B11], [B12], [B13].
尽管存在这种多样性,病理生理学机制却趋同于共同的神经生物学终点:兴奋-抑制(E/I)平衡失调、突触功能障碍、神经炎症、氧化应激以及肠脑轴的改变。 [B14], [B15], [B16].
兴奋/抑制(E/I)失衡假说认为,兴奋性(谷氨酸能)和抑制性(GABA能)神经传递之间的失衡可以解释自闭症谱系障碍(ASD)的核心症状。 B17, [B18].
影像学研究和死后分析显示GABA和谷氨酸浓度存在区域性改变,GABA-A和NMDA受体表达发生变化,以及GABA能中间神经元功能障碍。 [B19], [B20], [B21].
这种失衡尤其影响前额叶皮层(执行功能、社交认知)、杏仁核(情绪处理、社会感知)、小脑(感觉运动整合、认知过程)和海马体(记忆、情境加工)。 [B22], [B23], [B24].
神经炎症是另一个汇聚机制:死后研究显示自闭症谱系障碍(ASD)大脑中小胶质细胞和星形胶质细胞活化,临床前模型(例如,母体免疫活化、丙戊酸诱导的ASD)证明了促炎细胞因子(TNF-α、IL-6、IL-1β)的增加和慢性神经炎症导致的突触发育紊乱。 [B25], [B26], [B27].
已在多个自闭症谱系障碍(ASD)队列中发现氧化应激,其特征为活性氧(ROS)升高、谷胱甘肽(GSH)降低以及线粒体功能障碍。 [B28], [B29].
胃-脑轴的作用日益得到认可:自闭症患者常表现出肠道菌群失调,其厚壁菌/拟杆菌比例改变、微生物多样性减少以及神经递质前体(色氨酸、GABA)的产生受损。 [B30], [B31]. 临床前研究表明,粪便微生物移植可改善小鼠模型中的自闭症谱系障碍样行为,并调节血清素能和谷氨酸能信号通路。 [B32], [B33].
辅助疗法需求
尽管进行了深入研究,但仍没有针对自闭症谱系障碍核心症状的治愈性疗法。循证干预措施包括行为干预方法(应用行为分析、丹佛早期起步模式)和对症药物治疗。 [B34], [B35]. 利培酮和阿立哌唑是FDA批准的唯一用于ASD的药物,但仅限用于烦躁和攻击性行为,而非核心症状。 [B36], [B37]. 这些抗精神病药物与显著的副作用相关(体重增加、代谢综合征、锥体外系症状) [B38].
共病症状,如焦虑、睡眠障碍和感觉失调,通常用苯二氮䓬类药物、褪黑素或抗抑郁药治疗,但在自闭症谱系障碍人群中的有效性和安全性证据有限 [B39], [B40]. 这种对合并症症状缺乏安全有效疗法以及多重用药的负担,促使人们寻找辅助性的、副作用小的干预措施。 [B41].
精油及其主要成分(萜烯)作为一种潜在的辅助治疗选择,基于以下几点:
本报告分析了自闭症谱系障碍(ASD)的神经生物学基础、相关萜烯的分子作用机制、在ASD人群中的临床证据,并为精油作为多模式治疗方法的组成部分制定了基于证据的剂量和安全建议。.
自闭症谱系障碍的神经生物学
GABA/谷氨酸 E/I 不平衡
InEx/I不平衡假说是自闭症谱系障碍研究中最稳健的神经生物学概念之一 B17, [B18]. 谷氨酸是主要的兴奋性神经递质,GABA(γ-氨基丁酸)是主要的抑制性神经递质,它们共同调节神经元兴奋性、突触可塑性和网络动力学。兴奋性传递失衡(E/I比值升高)或抑制性控制受损会导致过度兴奋、感觉过滤受损以及社交信息处理能力下降。 [B45], [B46].
磁共振波谱(MRS)研究显示区域性改变:一些研究显示纹状体谷氨酸浓度降低,前额叶γ-氨基丁酸(GABA)水平升高,以及感觉和联合皮层中谷氨酸/GABA比值改变。 [B19], [B20].
验尸分析表明,在背外侧前额叶皮层和 cerebellum 中,GAD65 和 GAD67(GABA 合成酶)的表达降低,并且 GABA 能中间神经元(特别是 parvalbumin 阳性中间神经元)的密度发生改变。 [B47], [B48].
从发育生物学的角度来看,这是关键的:由于细胞内氯离子浓度高(由NKCC1共转运蛋白介导),GABA在早期发育阶段起兴奋性作用,并在出生后通过KCC2共转运蛋白的表达转变为抑制性作用。 [B49].
在 ASD 小鼠模型中已检测到 GABA 开关延迟或受损,并通过丁苯安 (NKCC1 抑制剂) 进行了纠正,从而改善了社交行为。 [B50], [B51].
一项使用布美他尼治疗自闭症谱系障碍儿童的临床试验显示,在社交沟通方面有中度改善,但存在方法学上的局限性。 B52.
谷氨酸能功能障碍的表现为NMDA和AMPA受体亚基表达改变、突触可塑性(长期增强/抑制)受损以及特定神经环路中谷氨酸能传递过度。 [B53], [B54].
遗传学研究已在谷氨酸能突触基因(GRIN2B、GRIA1、SLC1A1)中发现了与自闭症谱系障碍(ASD)相关的突变。 [B55].
临床前模型显示,调节谷氨酸能传递(例如通过mGluR5拮抗剂)可以改善ASD样行为 [B56].
血清素、多巴胺和催产素
单胺能系统在社会认知、奖赏处理和情绪调节中起着核心作用——这些都是自闭症谱系障碍(ASD)中受损的领域。 [B57], [B58].
羟色胺(5-HT)
高血清素血症(外周血清素水平升高)在 25%–50% 的 ASD 患者中被证实的,外周血清素与中枢 5-HT 水平的关系仍不清楚% [B59], [B60]. 影像学研究显示,自闭症儿童额叶皮层和丘脑的血清素合成能力降低 [B61]. 血清素转运蛋白 (5-HTTLPR) 和色氨酸羟化酶-2 (TPH2) 的遗传变异与自闭症谱系障碍 (ASD) 风险相关 [B62]. 临床前研究表明,在关键发育窗口期间的血清素能功能障碍会导致类似自闭症谱系障碍的表型。 [B63]. 选择性血清素再摄取抑制剂 (SSRIs) 在自闭症谱系障碍 (ASD) 中显示出喜忧参半的结果:对核心症状无效,对重复行为和焦虑有中度效果。 [B64].
多巴胺 (DA)
多巴胺能的边缘间和皮层间回路介导奖赏处理、动机和社交强化 [B65]. ASD 患者在面对社交奖励(如笑脸、社交互动)时,其腹侧纹状体的激活程度低于面对非社交奖励时。 [B66], [B67]. PET 研究报告显示纹状体多巴胺转运体密度增加,D2 受体可用性降低 [B68]. 遗传学研究确定了多巴胺能基因(DRD1、DRD2、DAT1)中与ASD相关的变异 [B69]. 临床前模型显示,多巴胺能功能障碍会导致社交偏好障碍和重复行为 [B70].
催产素(OT)
催产素是一种神经肽,在社会联系、信任、社会认知和压力调节中起着核心作用 [B71]. ASD 患者表现出降低的血浆催产素水平、催产素受体 (OXTR) 表达改变以及 OXTR 和 CD38(催产素释放)的基因变异。 [B72], [B73]. 鼻内催产素在一些研究中改善了自闭症成年人的社会认知和目光接触,但结果不一致,在儿科队列中无效。 [B74], [B75]. 临床前研究表明,催产素可调节GABA能发育:在关键时期给予催产素可纠正ASD小鼠模型中延迟的GABA转换并改善社交行为。 [B50], [B76]. 这些发现将氧促肾上腺素能和GABA能系统在机理上联系起来。.
神经炎症 – 小胶质细胞、TNF-α 和细胞因子
慢性神经炎症是自闭症谱系障碍(ASD)中一个汇聚性的病理生理学机制,这一观点得到了死后组织、影像学和生物标志物研究的支持。 [B77], [B78].
小胶质细胞激活
小胶质细胞是中枢神经系统的常驻免疫细胞,它们调节突触修剪、神经元发育和免疫监视 [B79]. 尸检研究显示,自闭症谱系障碍大脑的前额叶皮层、小脑和白质中存在激活的小胶质细胞(Iba1表达增加,变形虫形态)。 [B80], [b81]. PET成像与转运蛋白(TSPO)配体显示,在活体ASD患者的多个脑区中,小胶质细胞激活增加 [B82]. 活化的小胶质细胞释放促炎细胞因子,产生活性氧,并可以吞噬突触结构,从而导致连接紊乱。 [B83].
促炎细胞因子
在 ASD 患者的血浆、脑脊液和死后脑组织中检测到 TNF-α、IL-6、IL-1β 和 IFN-γ 水平升高。 [B84], [B85]. 母体免疫激活(MIA)模型,通过用聚(I:C)或LPS处理怀孕小鼠,可产生具有ASD样行为和TNF-α水平升高的后代 [B86], [B87]. .TNF-α 可调节突触传递、减少 GABA 能抑制并损害突触可塑性 B88. 阻断MIA模型中的TNF-α可预防ASD样表型 [B89].
星形胶质细胞功能障碍
星形胶质细胞通过谷氨酸转运蛋白 EAAT1/2 调节谷氨酸稳态、钾离子缓冲以及为神经元提供代谢支持 [B90]. 尸检研究显示自闭症大脑中存在星形胶质细胞增生(GFAP 表达升高) [B91]. 功能障碍的星形胶质细胞会引起细胞外谷氨酸的过度积聚和兴奋性毒性 [B92].
氧化应激和线粒体功能障碍
氧化应激源于 ROS 生成与抗氧化防御之间的失衡 [B93]. 多项研究报告称,自闭症谱系障碍 (ASD) 患者的氧化应激标志物升高:脂质过氧化(丙二醛)、蛋白质羰基化、DNA 氧化(8-OHdG)升高,以及抗氧化能力(谷胱甘肽、超氧化物歧化酶、过氧化氢酶)降低。 [B28], [B94], [B95].
谷胱甘肽耗竭
谷胱甘肽(GSH)是主要的细胞内抗氧化剂。ASD 患者血浆和大脑中的 GSH 水平降低,氧化型谷胱甘肽(GSSG)升高,GSH/GSSG 比值降低。 [B96], [B97]. 谷胱甘肽合成酶(GCLC、GSS)的遗传变异与自闭症谱系障碍(ASD)的风险有关 [B98].
线粒体功能障碍
5 至 30 % 的 ASD 患者显示出线粒体功能障碍的生化证据(乳酸升高、呼吸链活性降低) [B99], [B100]. 线粒体是 ROS 的主要来源;线粒体功能障碍导致 ROS 产生增加、ATP 合成减少和钙稳态紊乱 [B101]. .临床前研究表明,线粒体抗氧化剂(MitoQ)可改善小鼠模型中的类 ASD 行为 [B102].
Nrf2/HO-1 通路
核因子红细胞2相关因子2 (Nrf2) 是抗氧化反应的主调节因子,可诱导血红素加氧酶-1 (HO-1)、NAD(P)H:醌氧化还原酶-1 (NQO1)、谷胱甘肽S-转移酶等抗氧化酶的表达。 [B103]. 在ASD模型中已发现Nrf2活性降低;Nrf2激活剂(萝卜硫素)在小型临床试验中显示出ASD社会行为和沟通的改善。 [B104], [B105].
肠脑轴
胃肠道和中枢神经系统之间的双向沟通通过神经(迷走神经)、内分泌(下丘脑-垂体-肾上腺轴)、免疫和代谢途径进行 [B106]. ASD患者表现出高发的胃肠道症状(便秘、腹泻、腹痛)和菌群失调。 [B107], [B108].
微生物群落改变
荟萃分析显示,在自闭症谱系障碍(ASD)中,微生物多样性降低,厚壁菌门/拟杆菌门比例升高,双歧杆菌和普雷沃氏菌减少,梭菌和脱硫弧菌增加。 [B109], [B110]. 这些变化与症状的严重程度相关 [B111].
机械连接
肠道微生物群产生神经递质前体(色氨酸 → 5-羟色胺,谷氨酸 → GABA)、短链脂肪酸(丁酸盐、丙酸盐、乙酸盐)并调节免疫功能 [B112]. 丙酸酯暴露诱导啮齿动物模型中类似自闭症谱系障碍的行为 [B113]. 自闭症谱系障碍(ASD)患者的粪便微生物移植(FMT)到无菌小鼠会传播ASD样行为并改变大脑基因表达(血清素能、谷氨酸能突触基因)。 [B32]. .在 ASD 小鼠模型中对健康供体进行 FMT 可改善社交行为并使神经递质水平正常化 [B114].
迷走神经介导
迷走神经将信号从肠道传递到脑干(孤束核),并进一步传递到边缘系统和皮层区域。 [B115]. 临床前研究表明,电针通过迷走神经依赖性机制和微生物组调节改善了VPA小鼠模型中与ASD相似的行为;迷走神经切断术消除了这些影响 [B116].
突触功能障碍——神经细胞粘连分子、SHANK蛋白
突触功能障碍是自闭症谱系障碍的核心病理生理学概念,有遗传、分子和电生理学证据支持 [B117], [B118].
神经细胞粘附分子-神经细胞粘附蛋白复合物
神经粘素 (NLGN1-4) 是突触后细胞粘附分子,它们与突触前神经粘素 (NRXN1-3) 相互作用,并调节突触分化、成熟和功能。 [B119]. 在 ASD 患者中发现了 NLGN3 和 NLGN4X 的突变。 [B120]. NLGN3-R451C-基因敲除小鼠表现出抑制性传导增强、社交互动受损和重复行为 [B121]. NLGN4 基因敲除小鼠表现出兴奋性传递减弱和类似 ASD 的表型 [B122].
SHANK蛋白
SHANK1-3是突触后支架蛋白,它们组织谷氨酸受体、肌动蛋白细胞骨架和信号分子。 [B123]. .SHANK3 基因突变与 Phelan-McDermid 综合征(22q13 缺失)有关,该综合征的特征是自闭症、智力障碍和语言障碍。 [B124]. SHANK3基因敲除小鼠表现出树突棘密度降低、突触传递紊乱和类似自闭症谱系障碍的行为 [B125]. SHANK2 突变也与自闭症谱系障碍有关;SHANK2 基因敲除的小鼠表现出多动、刻板行为和 NMDA 受体功能障碍。 [B126].
突触可塑性
长时程增强(LTP)和长时程抑制(LTD)是学习和记忆的细胞学基础,在多种ASD小鼠模型中受到干扰 [B127]. 改变的LTP/LTD平衡可能导致经验依赖的电路成熟受损和社交学习受损 [B128].
受累的脑区
结构和功能成像在自闭症谱系障碍中识别了特定脑区的持续性改变 [B129], [B130].
前额叶皮层
前额叶皮层负责执行功能、工作记忆、认知灵活性和社会认知。 [B131]. ASD 患者在执行心理理论任务时表现出 PFC 激活减弱、与后部区域的 PFC 连接中断以及 PFC 微结构改变。 [B132], [B133]. 尸检研究显示,前额叶皮层(PFC)神经元密度增加、微柱宽度减小以及GABA能缺陷。 [B134].
杏仁核
杏仁核对于情绪处理、社会感知和恐惧条件反射至关重要。 [B135]. ASD儿童在儿童早期(2-4岁)表现出杏仁核增大,随后在青春期正常化或体积减小 [B136]. 功能性研究显示,在处理面部表情时,杏仁核的激活减弱,并且杏仁核与前额叶皮层(PFC)的连接受到干扰。 [B137], [B138].
小脑
小脑传统上与运动控制相关,但在认知过程、感觉预测和社会学习中也起着作用 [B139].
ASD 患者表现出浦肯野细胞丢失、小脑体积减小(尤其是小脑蚓部)以及小脑-皮层连接紊乱 [B140], [B141]. 小脑功能障碍可导致感觉预测受损、运动刻板以及社交时序处理受损。 [B142].
海马体
海马体介导陈述性记忆、空间导航和情境处理。 [B143]. ASD 患者在儿童时期海马体增大,海马体连接改变,以及情境性恐惧条件反射受损 [B144], [B145].
纹状体
纹状体(尾状核、壳核、伏隔核)介导奖赏处理、习惯学习和运动控制 [B146]. ASD患者表现出纹状体增大、社会奖励激活减少以及皮质纹状体连接受损。 [B147], [B148].
特异性神经发育障碍的代谢机制
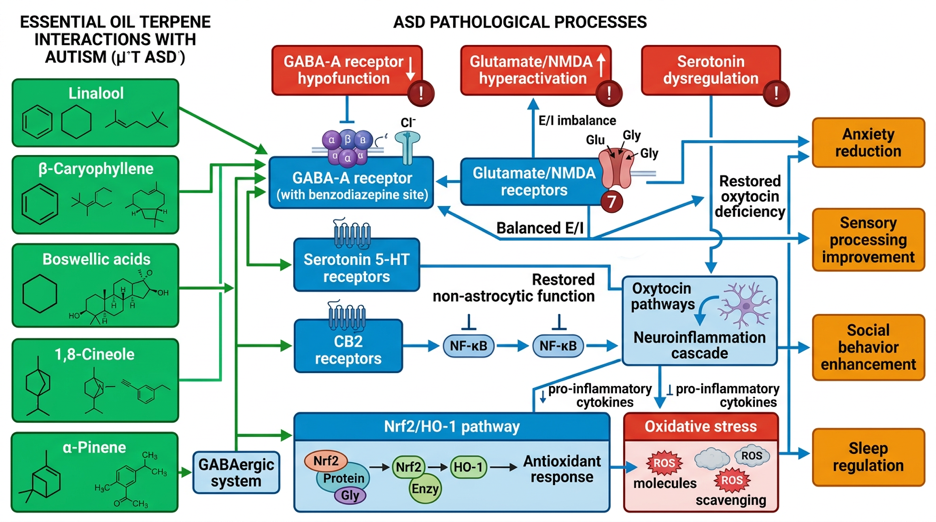
图 2:自闭症谱系障碍中萜类化合物的分子信号通路——GABA/谷氨酸平衡、神经炎症和氧化应激
萜烯是一类多样的天然产物,是精油的主要成分 [B149]. 其亲脂性使其能够穿过血脑屏障并与神经靶点相互作用 [B150]. 本节分析了参与 ASD 相关病理生理学靶点的特定萜烯的分子机制。.
芳樟醇 – GABA-A/苯二氮䓬受体、血清素、抗炎
芳樟醇(3,7-二甲基-1,6-辛二烯-3-醇)是一种单萜醇,也是薰衣草(狭叶薰衣草,25–45 %)、芫荽、佛手柑等精油的主要成分 [B151].
GABA-A受体调节
芳樟醇显示出抗焦虑作用,可通过氟马西尼(苯二氮䓬拮抗剂)阻断,这表明其与GABA-A受体的苯二氮䓬结合位点相互作用。 [B152]. 在一个嗅觉暴露模型中,芳樟醇蒸气在小鼠中产生了抗焦虑作用(高架十字迷宫、明暗箱),而在无嗅觉小鼠(嗅觉上皮损伤)中则不存在,这表明了嗅觉介导的作用。 [B152]. 电生理学研究表明,芳樟醇可增强培养神经元中GABA引起的氯化物电流。 [B153]. 这种GABA能调节对ASD高度相关,因为GABA能缺陷和兴奋/抑制(E/I)失衡是其核心病理生理机制。 B17, [B18].
血清素能调节
吸入芳樟醇可改变小鼠额叶皮层中血清素水平,提示血清素能传递受到调节 [B154]. 在一项针对健康成年人的研究中,有20分钟的芳樟醇吸入可以减轻抑郁情绪并增加副交感神经活动(提高心率变异性)。 [B155]. 离体研究表明,芳樟醇能提高下丘脑催产素神经元内钙浓度,这可能是一种催产素调节的机制。 [B155]. 这些发现将芳樟醇与 ASD 功能障碍的血清素能和催产素能系统联系起来。 [B59], [B72].
抗炎作用
系统性回顾报告了芳樟醇在临床前模型中的抗炎特性,包括肿瘤坏死因子-α (TNF-α)、白细胞介素-6 (IL-6)、白细胞介素-1β (IL-1β) 和核因子-κB (NF-κB) 激活的减少。 [B156]. 在应激诱导的神经炎症模型中,芳樟醇能减少小胶质细胞活化和促炎细胞因子表达。 [B157]. 这些抗神经炎症作用与自闭症谱系障碍(ASD)相关,因为慢性神经炎症是一种汇聚性的病理生理机制。 [B77], [B78].
药物动力学
吸入暴露后,芳樟醇迅速被吸收并进入体循环;代谢物(芳樟醇氧化物、葡萄糖醛酸苷)可在尿液中检测到。 [B158]. 由于脂溶性,血脑屏障的通过性很可能,但人体内直接的中枢神经系统药代动力学数据有限 [B159].
β-石竹烯 – CB2激动剂,NF-κB,神经炎症
β-石竹烯 (BCP) 是一种倍半萜碳氢化合物,存在于黑胡椒、丁香、大麻(非精神活性成分)、香脂油等植物中。 [B160].
CB2受体激动
BCP 是一种选择性大麻素 2 型受体(CB2)激动剂,对 CB1 受体无亲和力,可产生非精神活性的免疫调节作用 [B161]. CB2 受体主要在免疫细胞(小胶质细胞、外周免疫细胞)上表达,并调节细胞因子释放、小胶质细胞活化和神经炎症。 [B162]. 在CB2基因敲除小鼠中,BCP效应消失,证实了CB2的特异性。 [B163].
抗神经炎症作用
BCP 在多种临床前模型中可降低神经炎症:LPS 诱导的神经炎症、创伤性脑损伤、神经退行性疾病 [B164], [B165]. 机制包括减少小胶质细胞活化(Iba1、CD11b 表达降低)、抑制促炎细胞因子(TNF-α、IL-6、IL-1β)以及调节 NF-κB 信号通路。 [B166]. 在母体免疫激活模型(MIA,与ASD相关的模型)中,BCP治疗改善了ASD样行为(社交缺陷、重复行为),并减少了后代的神经炎症。 [B167].
GABA能和肾上腺素能调节
行为药理学研究表明,BCP的抗焦虑和抗抑郁作用会被氟马西尼(苯二氮䓬受体拮抗剂)和L-NAME(一氧化氮合酶抑制剂)阻断,这提示GABA能和硝能系统参与其中。 [B168]. 这些发现表明 BCP 可能通过 CB2 介导的神经炎症减轻间接调节 GABA 能功能。.
抗氧化作用
BCP 主要通过减少活性氧(ROS)和脂质过氧化,以及增强内源性抗氧化酶(SOD、过氧化氢酶、GSH)的活性来展现抗氧化特性。 [B169]. 在一个氧化应激模型中,BCP 保护神经元免受 H₂O₂ 诱导的细胞凋亡 [B170].
ASD的临床相关性
CB2介导的神经炎症减轻、抗氧化作用和间接GABA能调节的结合,使得BCP成为一种在机理上具有合理性的ASD辅助治疗候选药物,特别是对于具有神经炎症特征的患者。 [B171].
乳香酸 – Nrf2/HO-1,抗神经炎症
乳香酸是从乳香树种(乳香)的树脂中提取的五环三萜酸,包括β-乳香酸、11-酮基-β-乳香酸(KBA)和3-O-乙酰基-11-酮基-β-乳香酸(AKBA) [B172].
Nrf2/HO-1 激活
乳香酸激活 Nrf2 信号通路,这是抗氧化反应的主调节因子 [B173]. 在小鼠的神经炎症模型中,乳香提取物治疗提高了海马体和皮层中 Nrf2 的核定位和 HO-1 的表达 [B174]. HO-1催化血红素分解为胆绿素(抗氧化)、一氧化碳(抗炎、血管扩张剂)和铁;HO-1诱导可预防氧化应激和神经炎症。 [B175].
抗神经炎症机制
乳香酸可降低小胶质细胞和星形胶质细胞的活化(降低 Iba1、GFAP 表达),抑制促炎细胞因子(TNF-α、IL-6、IL-1β),并抑制 NF-κB 和 MAPK 信号通路。 [B176], [B177]. 在 LPS 诱导的神经炎症模型中,乳香治疗改善了认知障碍并降低了神经炎症标志物 [B178].
5-脂氧合酶 (5-LOX)-抑制
乳香酸,特别是AKBA,可抑制5-LOX,这是一种将花生四烯酸代谢为促炎性白三烯的酶 [B179]. 5-LOX 抑制可减少白三烯介导的炎症,并在临床前研究中与神经保护作用相关。 [B180]. 然而,现有文献中关于勃拉沃素作为减轻脑损伤的主要机制的证据有限;Nrf2/HO-1 激活似乎是主要机制 [B174].
自闭症谱系障碍相关模型的临床前证据
在丙戊酸(VPA)诱导的自闭症谱系障碍(ASD)小鼠模型中,乳香提取物治疗改善了社交互动,减少了重复行为,并使神经炎症标志物正常化。 [B181]. 这些发现支持了这样一个假设,即乳香介导的神经炎症减轻和抗氧化作用可以改善与自闭症相关的行为。.
1,8-桉叶油醇 – GABA-A/苯二氮䓬类,抗焦虑
1,8-桉叶素(桉叶油素)是一种单萜氧化物,是桉树(蓝桉,70-90 %)、迷迭香、茶树和月桂的主要成分。 [B182].
GABA-A受体调节
吸入1,8-桉油醇在小鼠中产生了剂量依赖性的抗焦虑和抗抑郁作用(高架十字迷宫、强迫游泳试验、悬尾试验) [B183]. 这些效应被氟马西尼阻断,这表明它们与GABA-A受体的苯二氮䓬结合位点发生了相互作用。 [B183]. 已证明存在剂量依赖性:低剂量(0.1–1 µg/小鼠,吸入)显示出抗焦虑作用,而高剂量(10 µg/小鼠)则产生镇静作用 [B184].
抗炎和抗氧化作用
1,8-桉油醇通过抑制NF-κB、降低TNF-α、IL-1β和白三烯的合成而表现出抗炎特性 [B185]. 在LPS诱导的神经炎症模型中,1,8-桉树脑减少了小胶质细胞活化和氧化应激 [B186].
安全方面
由于存在喉痉挛和呼吸抑制的风险,3 岁以下儿童禁用 1,8-Cineole [B187]. 在正确使用(吸入、低剂量)的情况下,1,8-桉树脑对成人和较大儿童的安全性良好 [B188].
α-蒎烯 – GABA能
α-松油烯是一种双环单萜,存在于松树、迷迭香、鼠尾草和大麻中 [B189].
GABA能调节
系统性回顾将 α-蒎烯与 GABA 能神经递质的调节相关联,尽管直接的作用机制研究有限 [B190]. 在行为模型中,α-蒎烯表现出抗焦虑和镇静作用,提示其作用涉及GABA能机制。 [B191]. 现有文献中缺乏关于直接GABA-A受体相互作用的电生理学数据。.
抗炎和神经保护作用
α-蒎烯通过抑制 NF-κB 和减少促炎细胞因子而表现出抗炎特性 [B192]. 在一个缺血再灌注模型中,α-蒎烯可预防神经损伤并减轻氧化应激 [B193].
乙酰胆碱酯酶 (AChE) 抑制
一些研究报告称α-蒎烯具有乙酰胆碱酯酶抑制活性,这可能增强胆碱能神经传递 [B194]. 然而,其与ASD的相关性尚不清楚,因为胆碱能功能障碍并非ASD的主要病理生理机制。 [B195].
依兰/香叶- 5-羟色胺/多巴胺在VPA模型中
依兰(香叶卡纳加)精油含有芳樟醇、香叶醇、β-石竹烯等萜烯 [B196].
VPA-ASD模型中的临床前证据
依兰依兰精油吸入改善了丙戊酸(VPA)诱导的自闭症谱系障碍(ASD)大鼠的类焦虑行为、社交互动和认知功能 [B197]. 神经化学分析表明,依兰(Cananga odorata)能够调节前额叶皮层和海马体的血清素和多巴胺代谢:血清素水平升高,5-HIAA/血清素比例降低(表明血清素周转率降低),多巴胺水平正常化。 [B197].
机械论解释
通过调节血清素能和多巴胺能系统来改善自闭症谱系障碍(ASD)样行为在机理上是合理的,因为这两种系统在自闭症谱系障碍(ASD)中都存在功能障碍。 [B57], [B58]. 尚未确定依兰依兰中导致这些作用的特定成分;基于其已知机制,芳樟醇和 β-石竹烯是可能的候选物 [B152], [B161].
局限性
这些发现来自一个单一的临床前模型(VPA大鼠);在其他ASD模型中的重复以及向人类研究的转化尚不充分。 [B197].
ASD 中特定的精油和临床证据
薰衣草/芳樟醇 – ABA 合规性、催产素神经元
ASD的临床证据
一项小型试点研究(n = 12 名 ASD 儿童)研究了薰衣草芳香疗法作为应用行为分析 (ABA) 疗法辅助手段。 [B198]. 薰衣草在治疗期间以“20滴加入100毫升”(可能是水或载体油,细节不清楚)的比例进行扩散。研究报告称,与对照组的治疗相比,薰衣草暴露期间的依从性得分(合作、任务完成)更高。 [B198]. 局限性包括样本量非常小、缺乏随机化、盲法不明确以及缺乏标准化剂量。.
一项对四名患有严重学习障碍的孤独症儿童进行的观察性研究,将芳香疗法按摩(薰衣草、天竺葵、橘子和载体油)融入日常作息。 [B199]. 研究报告称,在引入芳香疗法按摩后,共享注意力行为(眼神交流、共同活动)有所增加。 [B199]. 局限性包括缺乏控制、样本量小以及由按摩(触觉刺激、社交互动)效果引起的混淆。.
机械学基础
临床发现与临床前机制一致:芳樟醇介导的GABA能调节可降低焦虑和改善合作;血清素能和催产素能调节可促进社交注意力和互动 [B152], [B155]. 一项针对健康成年人的研究表明,吸入薰衣草可减轻抑郁情绪,并在体外实验中提高了下丘脑催产素神经元细胞内钙的浓度 [B155]. 这种氧促褪黑素的调节与自闭症谱系障碍高度相关,因为催产素功能障碍是一个已确立的病理生理机制。 [B72].
证据的局限性
在自闭症谱系障碍(ASD)方面,薰衣草的临床证据质量非常低:样本量小、缺乏随机化、掩蔽不足、干预措施不统一且缺乏重复性。 [B198], [B199]. 缺乏使用标准化薰衣草制剂、客观结果测量和足够功效的、高质量的随机对照研究。.
佛手柑 – ASD 儿童的随机对照试验,结果不显著
随机对照试验
一项随机、双盲研究探讨了在使用芳香疗法(芳香精油)时,对 ASD 儿童(n=42)的药物诱发性焦虑的影响 [B200]. 儿童被随机分为15分钟吸入佛手柑油或安慰剂(水)后进行医疗检查。主要结局指标是通过改良耶鲁术前焦虑量表(mYPAS)测量的焦虑。在调整了基线焦虑后,佛手柑组与安慰剂组相比,焦虑水平没有显著降低(p > 0.05) [B200].
口译
这是唯一确定的关于自闭症儿童使用精油的随机对照研究。负面结果可能有多种原因:(1) 暴露时间短(15分钟)可能不足够;(2) 医疗环境下的急性焦虑可能比慢性焦虑更难调节;(3) 嗅觉敏感性和偏好存在个体差异;(4) 原本基线焦虑水平较高可能存在的封顶效应。 [B200].
临床前机制
临床前研究表明,佛手柑在啮齿类动物模型中具有抗焦虑作用。 [B201]. 有趣的是,这些效果并未被氟马西尼阻断,这表明存在非苯二氮䓬类介导的机制,并将佛手柑与芳樟醇和桉树脑区分开来。 [B201]. 佛手柑含有芳樟醇、乙酸芳樟酯、柠檬烯和佛手柑内酯;具体负责抗焦虑作用的成分尚不清楚。 [B202].
临床影响
这项随机对照试验(RCT)的阴性结果,降低了将佛手柑用于自闭症谱系障碍(ASD)急性焦虑的预期。在推荐佛手柑用于ASD之前,需要进行更长时间暴露、慢性焦虑结局和其他ASD人群的进一步研究。 [B200].
乳香 / 锯叶乳香 - 抗神经炎症,Nrf2
ASD的临床证据
未发现针对自闭症谱系障碍(ASD)患者使用乳香(Boswellia)精油或乳香提取物的随机对照研究。现有文献不包含乳香针对ASD的临床数据。 [B203].
临床前证据
如第 3.3 节所述,乳香酸在临床前模型中显示出强大的抗神经炎症和抗氧化作用,包括 Nrf2/HO-1 激活、小胶质细胞抑制以及在 VPA 小鼠模型中改善 ASD 样行为。 [B174], [B181]. 这种机制上的合理性支持了 Boswellia 可能对具有神经炎症特征的 ASD 患者有益的假设。.
其他适应症的临床应用
乳香提取物(口服)在临床上用于治疗炎症性疾病(关节炎、哮喘、炎症性肠病),疗效证据中等,安全性良好。 [B204]. 吸入焚香精油是传统的用法,但临床药代动力学和疗效数据有所缺乏。 [B205].
研究需求
需要进行使用勃林卡(口服或吸入)治疗自闭症谱系障碍(ASD)患者的对照临床试验,最好按照神经炎症生物标志物进行分层,以评估临床疗效。 [B206].
雪松木/香柏木 – 镇静,GABA能
临床前和临床证据
现有文献未包含有关雪松木(香柏属、弗吉尼亚刺柏)精油在自闭症谱系障碍(ASD)或自闭症谱系障碍相关临床前模型中的具体机制或临床数据 [B207]. 传统应用包括镇静和焦虑缓解,但分析文献中缺乏 GABA 能机制或临床疗效的科学证据 [B208].
主要成分
雪松油含有雪松醇(倍半萜烯醇)、α-雪松烯和侧柏酮。 [B209]. 雪松醇与镇静特性有关,但缺乏关于GABA-A受体相互作用或神经炎症调节的直接研究 [B210].
结论
迄今为止,关于雪松木在自闭症谱系障碍(ASD)治疗中的证据不足,无法根据现有文献提出相关建议。 [B207].
香根草 – 注意
临床前和临床证据
现有文献未包含关于 ASD 或 ASD 相关模型中香根草(香根草)精油的具体数据 [B211]. 轶事报道和传统应用表明了对注意力和多动症的影响,但缺乏对照研究 [B212].
主要成分
香根草油含有香根草醇、香根草酮、α-香根酮和β-香根酮(倍半萜醇类和酮类)。 [B213]. 在已分析的文献中,缺乏关于这些组分在神经递质系统或神经炎症方面的机制研究。 [B214].
结论
自闭症谱系障碍(ASD)中香根草的证据不足;在提出建议之前需要进行进一步的研究。 [B211].
依兰依兰 - 血清素/多巴胺,VPA模型
临床前证据
如第3.5节所述,依兰(Cananga odorata)精油吸入改善了VPA大鼠的ASD样行为,并调节了前额叶皮层和海马的血清素和多巴胺代谢。 [B197]. 这是唯一一项在 ASD 相关模型中进行依兰依兰研究的已识别研究。.
ASD的临床证据
未发现使用依兰依兰治疗自闭症谱系障碍患者的临床试验。 [B215].
其他适应症的临床应用
依兰依兰传统上用于治疗焦虑、改善情绪和放松。一项针对健康成年人的小型研究表明,吸入依兰依兰可降低血压和心率,并提高主观的平静感。 [B216]. 关于血清素能和多巴胺能对人类影响的机制研究的中文翻译是:关于血清素能和多巴胺能对人类影响的机制研究尚付阙如。 [B217].
结论
临床前证据很有希望,但尚未转化为人类自闭症谱系障碍研究。依兰依兰可能被考虑作为人类自闭症谱系障碍临床试验的候选药物,特别是针对焦虑和情绪症状。 [B197].
ASD 的其他佐证精油
根据最新的文献研究,还识别出以下附加的精油,它们可能通过神经生物学上合理的机制对自闭症谱系障碍(ASD)产生影响。.
姜 (Zingiber officinale) – AKT/GSK3β 信号通路
活性成分
6-姜酚
6-姜辣素
姜烯
β-倍半萜苯
受体和作用机制
AKT/GSK3β信号通路:磷酸化增加→突触可塑性增强
抗神经炎症:抑制STAT3磷酸化和TNF-α
神经发生:临床前模型中海马神经发生标志物的改善
- 胶质细胞抑制:减少反应性星形胶质细胞增生
临床/临床前证据
- 临床前研究(VPA-小鼠模型)
– 口服姜提取物(4周,从第6周开始)显著改善了社交互动,减少了类似焦虑的行为,并提高了记忆表现 [B347]
– 机制:AKT/GSK3β 上调,STAT3/TNF-α 抑制,突触形成改善
尽管自闭症谱系障碍(ASD)的临床随机对照试验(RCT)仍然缺乏,但临床前证据强劲且具有良好的机理基础。
用量(Organik Aromas Nebuliser 3.0)
– 2–3 滴,适用于 15 至 25 平方米的房间
– 与薰衣草(1:2)混合,具有放松效果
– 用途:社交活动前 30 分钟
安全
易于耐受;3 岁以下儿童慎用。.
檀香(Santalum album)– α/β-檀香醇
活性成分
α-檀香醇(50-55%)
β-檀香醇(20–25%)
倍半萜
受体和作用机制
血清素能调节:α-檀香酚调节 5-HT 受体(临床前)
抗焦虑:在动物模型中已记录 GABA 能活性
镇静/舒缓:适用于自闭症谱系障碍的感官过敏
临床证据
檀香木是 Hiwa 糖浆 RCT(波斯医学,多种草药)用于 ASD 儿童的组成部分,其效果不能单独归因。 [B348]
– 单项研究表明,在健康受试者中,抗焦虑药物具有镇静作用
– 缺乏直接的 ASD 特定研究,仅使用纯檀香油
用量(Organik Aromas Nebuliser 3.0)
– 3-4 滴,适用于 15 至 25 平方米的空间
– 特别适合有睡眠困难的 ASD 儿童的睡前/助眠仪式
- 建议与薰衣草 (1:1) 混合使用
安全
耐受性极佳;最安全的精油之一,适用于儿童(2岁以上)。.
萜烯混合物(受大麻启发)——亲社会效应
活性成分
月桂烯
酸橙
芳樟醇
松子
β-石竹烯(作为混合物)
受体和作用机制
CB1/CB2-激动剂:萜烯混合物增强CBD效果(随行效应)
GABA 能调节:芳樟醇 + 氢化月见草协同作用
CBD 以外的亲社会效应已得到证实
临床证据
- BTBR 小鼠模型(ASD 模型)
吸入萜烯混合物(月桂烯、柠檬烯、芳樟醇、蒎烯、β-石竹烯)显示出独立于 CBD 的急性亲社会效应 [B349] - 萜烯混合物在联合应用时增强了 CBD 的作用
– 提示旁观者效应作为自闭症谱系障碍的治疗原则
用量(Organik Aromas Nebuliser 3.0)
- 混合建议:薰衣草(2 茶匙)+ 黑胡椒(1 茶匙)+ 佛手柑(1 茶匙)+ 迷迭香(1 茶匙)
– 15号房间是25平方米:总量5滴
- 用法:社交活动或团体治疗前 30 分钟
Organik Aromas Nebulizer 3.0 的剂量建议
Der Organik Aromas Nebulizer 3.0 采用文丘里管原理(伯努利效应)进行无水冷雾化精油 [B218]. 与稀释或加热油的超声波或热扩散器不同,喷雾器可产生纯精油的细小气溶胶颗粒(1至5微米),而不会发生热降解。 [B219]. 这使得室内空气中的挥发性萜烯浓度更高,并可能产生更强的药理作用 [B220].
文丘里效应与冷雾化
文丘里原理描述了流体在流经收缩段时压力的降低 [B221]. 在雾化器中,压缩空气通过一个喷嘴,产生负压,将精油从储罐中吸出并雾化成细小的液滴。 [B222]. 这些液滴被释放到室内空气中,在那里被吸入或沉降到表面上 [B223].
冷雾化优点
- 热敏成分的获取(无热降解) [B224]
– 挥发性萜烯浓度更高(无水分稀释效应) [B225]
极细粒径(1至5微米)可实现深层呼吸道穿透 [B226]
缺点
与超声波扩散器相比,成本更高 [B227]
– 较高的机油消耗 [B228]
– 潜在的过量风险,如果使用不当 [B229]
按房间大小划分的滴水量
剂量建议基于空间体积、换气率、期望的萜烯浓度和暴露持续时间。以下建议是保守的,并考虑了 ASD 特定的感觉过敏。 [B230].
表 1:Organik Aromas Nebulizer 3.0 针对不同空间大小的剂量建议
| 房间面积 (平方米) | 空间体积(立方米,层高 2.5 米) | 滴精油 | 使用时间 (分钟) | 暂停(分钟) | 日常应用 |
| 10–20 | 25–50 | 2–4 | 15–20 | 30–60 | 2–3 |
| 20–40 | 50–100 | 4–6 | 20–30 | 30–60 | 2–3 |
| 40–60 | 100–150 | 6–10 | 30 | 60 | 2 |
| >60 | 150 | 10–15 | 30 | 60 | 1–2 |
说明
- 滴定管容积: 1滴 ≈ 0,05毫升精油 [B231]
- 起始剂量 从最低推荐的滴数 [B232]
- 个性化定制 ASD 患者的嗅觉过敏可能需要较低的剂量或更长的休息时间 [B233]
- 通风 良好的通风对于避免过度积聚至关重要 [B234]
使用时间和休息时间
使用持续时间
基于临床前吸入研究,每次会议 15 至 30 分钟足以达到治疗性萜烯浓度 [B183], [B184]. 较长时间(>30分钟)的使用会增加感觉超负荷和嗅觉适应(持续暴露后感知能力下降)的风险 [B235].
暂停
30 至 60 分钟的应用间隔可实现嗅觉恢复、吸入萜烯的代谢以及室内空气浓度的降低 [B236]. 持续暴露而不休息可能会导致头痛、恶心和感官厌恶,特别是对于患有感官过敏症的 ASD 患者。 [B237].
日常应用
每天 2 到 3 次(早、中、晚)的用量对大多数适应症来说都足够了 [B238]. 每天超过4次的使用会增加副作用的风险,而不会带来额外的治疗益处。 [B239].
按时段划分的协议
早上 (提神,专注)
- 油 迷迭香 (1,8-桉树脑、α-蒎烯), 薄荷 (薄荷醇、薄荷酮), 柠檬 (柠檬烯), 香柠檬 (芳樟醇、乙酸芳樟酯)
- 用量: 3–5 滴 适用 20–30 平方米 房间
- 持续时间 15–20 分钟
- 目标: 促进学校/治疗期间的警觉性、注意力及认知功能 [B240]
下午(有助镇定,调节情绪)
- 油 佛手柑,依兰,天竺葵,乳香
- 用量: 3–5 滴 适用 20–30 平方米 房间
- 持续时间 20–30分钟
- 目标: 减少下午的焦虑,稳定情绪,活动间的过渡 [B241]
晚上(舒缓,助眠)
- 油 薰衣草 (芳樟醇、乙酸芳樟酯)、罗马洋甘菊 (酯类)、雪松 (雪松醇)、香根草
- 用量: 4-6滴,适用于20-30平方米的房间
- 持续时间 睡前三十分钟
- 目标: 减少晚间焦虑,促进放松和入睡 [B242]
ASD 特征
感觉过敏 60–90 % der ASD-Patienten zeigen olfaktorische Überempfindlichkeit oder Aversion gegenüber bestimmten Gerüchen [B243]. 渐进式介绍是必不可少的:
- 第一周: 室外展览(在相邻房间中扩散的油)以评估接受度 [B244]
- 第二周: 最低剂量(2滴),喷雾10分钟,喷雾期间患者离开房间 [B245]
- 第三周: 最低剂量,病人已在室内,观察耐受迹象(放松,无厌恶感) [B246]
- 第四周+ 根据个体耐受性进行的剂量和持续时间的渐进式增加 [B247]
基于偏好的选择
让患者(如有可能)闻不同的香油,并表达偏好。嗅觉偏好与治疗接受度和依从性相关。 [B248].
视觉和听觉线索
使用视觉计时器或听觉信号来指示芳香疗法疗程的开始和结束,这增加了自闭症谱系障碍患者的可预测性和控制感 [B249].
家长/监护人培训
对家长和看护者进行培训,使其能够识别容忍的迹象(放松、注意力改善)和厌恶的迹象(皱眉、退缩、烦躁)。 [B250].
质量要求 – GC/MS 分析
气相色谱-质谱联用(GC/MS)
GC/MS是精油化学表征的金标准 [B251]. GC 根据沸点和极性分离挥发性组分;MS 根据质荷比鉴定组分 [B252]. 气相色谱-质谱联用分析可实现:
- 主成分识别: 对具有治疗作用的萜烯(例如薰衣草中的芳樟醇、桉树中的1,8-桉树脑)的存在进行确认 [B253]
- 量化 各组分百分比组成 [B254]
- 欺诈检测 合成添加剂、稀释剂或污染物的识别 [B255]
- 批次到批次一致性: 确保可重复的组成 [B256]
治疗用精油的质量标准
- 100 % 莱茵
无合成添加剂、载体油或稀释剂 [B257] - 治疗级
在最佳条件下种植、收获和蒸馏的植物油 [B258] - Organisch/Wildsammlung
不含杀虫剂、除草剂和化肥 [B259] - GC/MS 证书
每批次都应附带 GC/MS 分析证书,其中应包含主要成分和纯度信息。 [B260] - 植物学名
植物物种的明确识别(例如,狭叶薰衣草,而非仅“薰衣草”) [B261] - 原产国和蒸馏日期
透明性有关溯源和新鲜度 [B262]
储存
精油应储存在深色玻璃瓶中,并置于阴凉处(<25°C),以最大程度地减少氧化和降解。 [B263]. 氧化后的油可能会引起皮肤刺激,并降低治疗效果 [B264].
安全说明和禁忌症
ASD 特异性感觉超敏
感觉处理障碍存在于 60%–90% % 的 ASD 患者中,表现为对感觉刺激的超敏(过度敏感)或低敏(不敏感)。 [B265]. 嗅觉过敏可能导致厌恶、焦虑、恶心和回避行为 B266.
风险管理
- 毕业介绍 如5.4节所述,从患者房间外开始使用最低剂量 [B244]
- 偏好筛选 在治疗应用前测试油以识别不良反应 [B248]
- 终止标准 一旦出现痛苦迹象(烦躁、退缩、恶心、头痛),请立即停止 [B267]
- 替代应用途径: 对于吸入性不耐受,请考虑局部使用(用载体油稀释,需进行斑贴试验)或被动扩散(将精油滴在房间角落的棉垫上)。 [B268]
癫痫风险——含樟脑的精油
10–30 % der ASD-Patienten haben komorbide Epilepsie [B269]. 某些精油,特别是含樟脑的精油,会降低惊厥阈值并可能引发癫痫发作 [B270].
癫痫禁忌的油
- 樟树 (Cinnamomum camphora): 包含樟脑(含量高达 50 %),一种已知的惊厥剂 [B271]
- 迷迭香(迷迭香,樟脑化学型): 含有 15-25 % 樟脑 [B272]
- 鼠尾草(Salvia officinalis): 含有侧柏酮(10–40 %),一种具有惊厥特性的GABA-A拮抗剂 [B273]
- 牛至 (Hyssopus officinalis): 含有可能引起癫痫发作的松香脑和异松香脑 [B274]
- 茴香 (Foeniculum vulgare): 包含反式茴香脑,高剂量可能引起抽搐 [B275]
癫痫的安全替代疗法: 薰衣草(狭叶薰衣草,芳樟醇化学型):临床前研究中的抗惊厥特性 [B276] 罗马洋甘菊(Anthemis nobilis):未发现促癫痫发作作用 [B277] – 香柠檬(香柠檬,无佛手柑内酯):对癫痫安全 [B278]
临床建议: 对于患有癫痫的ASD患者,应仅使用不含樟脑或侧柏酮成分的精油。建议在引入芳香疗法前咨询神经科医生。 [B279].
3岁以下儿童的桉树
1,8-桉叶油脑(桉叶油的主要成分)在3岁以下儿童中禁用,因为有喉痉挛、支气管痉挛和呼吸抑制的风险。 [B280]. 机制包括对呼吸道粘膜的直接刺激和迷走神经反射的触发 [B281].
3岁以下儿童禁忌用油
– 桉树(蓝桉,辐射松):70–90 % 1,8-桉叶素 [B282]
迷迭香(迷迭香,1,8-桉叶素-化学型):40–50 % 1,8-桉叶素 [B283]
– 茶树 (互叶白千层) (Melaleuca alternifolia): 5–15 % 1,8-桉叶素 (风险较低,但仍需谨慎) [B284]
- 薄荷 (Mentha piperita):薄荷醇可能引起类似的呼吸道刺激 [B285]
3岁以下儿童的安全替代品
薰衣草(狭叶薰衣草):在正确稀释的情况下,从出生起就安全 [B286]
罗马洋甘菊 (Anthemis nobilis): 从出生起即可安全使用 [B287]
– 橘子 (Citrus reticulata):6个月以上安全 [B288]
临床指南
3岁以下的ASD儿童应避免使用富含1,8-桉树脑的精油。.
对于 3–6 岁的儿童,请使用低剂量(成人剂量的 50%),并密切观察呼吸道症状。 [B289].
抗精神病药物相互作用:利培酮、阿立哌唑
利培酮和阿立哌唑是FDA唯一批准的用于自闭症谱系障碍(ASD)易怒的药物 [B290]. 两种药物都通过细胞色素P450酶(CYP2D6,CYP3A4)代谢 [B291]. 一些精油和萜烯可以抑制或诱导CYP酶,这可能导致药物水平改变。 [B292].
潜在的相互作用:
佛手柑
含有香柠檬内酯(呋喃香豆素),可抑制CYP3A4 [B293]. 理论上,香柠檬可能会增加瑞斯哌酮和阿立哌唑的水平,这可能会加剧副作用(镇静、锥体外系症状)。 [B294]. 然而,吸入暴露的水平远低于口服摄入(西柚汁效应);吸入佛手柑的临床相关性尚不清楚 [B295].
建议
对使用利培酮/阿立哌唑的患者使用不含佛手柑香豆素的佛手柑 (FCF,呋喃香豆素游离) [B296].
葡萄柚
葡萄柚精油含有呋喃香豆素并抑制CYP3A4 [B297]. 避免在接受CYP3A4代謝藥物的患者中使用 [B298].
圣约翰草(贯叶连翘)
虽然圣约翰草(贯叶连翘)不是一种典型的精油,但在芳香疗法的语境中偶尔会被提及。圣约翰草(贯叶连翘)是一种强效的CYP3A4诱导剂,可能会降低利培酮/阿立哌唑的水平,从而导致疗效下降。 [B299]
禁忌症 在服用抗精神病药物的患者中避免使用圣约翰草 [B300].
薰衣草, 罗马洋甘菊, 雪松
未发现已知的临床相关的CYP相互作用 [B301]. 在瑞斯匹瑞酮/阿立哌唑患者中安全 [B302].
药效学相互作用
镇静精油(薰衣草、洋甘菊、雪松)可能对精神科药物产生累加的镇静作用 [B303]. 这可能是治疗上可取的(例如助眠),但需要监测过度镇静。 [B304].
临床指南
告知处方医生有关芳香疗法的应用。对接受CYP3A4底物治疗的患者使用不含香柠檬素的精油。监测药物作用的变化(镇静、副作用)。 [B305].
质量要求:GC/MS 分析
正如第 5.5 节所述,GC/MS 分析对于确保纯度、成分和没有污染物至关重要 [B251]. 掺假或受污染的精油可能引起过敏、皮肤刺激、呼吸道症状和不可预测的药理作用。 [B306].
常见的造假
- 合成添加剂 合成芳樟醇或乙酸芳樟酯在“薰衣草”油中 [B307]
- 稀释剂 丙二醇, 二丙二醇甲醚 (DPM) [B308]
- 更便宜的油 薰衣草(Lavandula x intermedia)被当作真正薰衣草(Lavandula angustifolia)出售 [B309]
- 杀虫剂: 非有机油中的农药、除草剂残留 [B310]
安全建议
仅使用 GC/MS 认证的、100 % 纯净的有机精油,并确保其来源可靠。要求提供每批次的 GC/MS 认证。 [B311].
讨论
证据基础
关于精油对自闭症谱系障碍 (ASD) 的证据不尽相同,且大多质量较低。临床文献包括一项阴性的随机对照试验(佛手柑对急性焦虑) [B200], 小规模的试点研究存在方法上的局限性(薰衣草与 ABA 合规性) [B198], 观察性研究(芳香疗法按摩) [B199] 和病例报告 [B312]. 尚无研究能以足够证据质量证明其对自闭症核心症状(社交沟通、重复行为)的疗效 [B313].
与之形成对比的是,临床前证据充分:多项研究表明,特定的萜烯(芳樟醇、β-石竹烯、乳香酸、1,8-桉树脑)能够靶向 与ASD相关的分子靶点(GABA-A受体、CB2受体、Nrf2通路、神经炎症),并改善动物模型中类似ASD的行为。 [B152], [B161], [B174], [B183], [B197]. 这种临床前合理性与临床证据之间的差异在补充疗法中很常见,并且反映了研究资金的不足、方法上的挑战以及转化差距。 [B314].
机械合理性
基于自闭症谱系障碍(ASD)的病理生理学与萜烯的作用机制之间的趋同性,对精油治疗ASD的机制合理性是存在的:
GABA/谷氨酸 E/I 不平衡
芳樟醇和1,8-桉树脑调控GABA-A受体在苯二氮䓬结合位点,这可能增强抑制性神经传递并使E/I平衡正常化 [B152], [B183]. 这些机制类似于布美他尼,布美他尼在临床研究中显示出对ASD有温和的改善。 B52.
神经炎症
β-石竹烯(CB2激动剂)和乳香酸(Nrf2激活剂)在临床前模型中可减少小胶质细胞活化、促炎细胞因子和氧化应激 [B161], [B174]. 由于神经炎症是自闭症谱系障碍中一个既定的病理生理机制 [B77], 这些萜烯是否可能对患有神经炎症特征(外周细胞因子升高,PET显示小胶质细胞活化)的患者有用 [B315].
血清素/多巴胺
在临床前研究中,芳樟醇和依兰依兰调节血清素能和多巴胺能系统 [B154], [B197]. 这些机制可能会改善共病症状(焦虑、情绪),尽管不太可能对核心症状产生影响 [B316].
催产素
芳樟醇可增加催产素神经元中细胞内钙浓度 [B155]. 催产素调节 GABA 能发育,并在临床前研究中改善了类似 ASD 的表型 [B50], 芳樟醇是否可能间接调节氧化型多巴胺能信号通路 [B317].
肠脑轴
尽管缺乏直接证据,但抗炎萜类化合物(β-石竹烯、乳香酸)可能通过间接调节肠脑轴信号通路来减少全身炎症。 [B318]. 这项假说需要进行实验验证 [B319].
局限性
小型研究
大多数自闭症谱系障碍特异性研究的样本量小于 20,这意味着它们的效力不足以检测出中等程度的影响。 [B320]. 由于异质性,无法进行荟萃分析 [B321].
没有随机对照试验来治疗核心障碍
尚无关于评估精油对ASD核心症状(社交沟通、重复性行为)疗效的随机对照研究。 [B322]. 唯一确定的随机对照试验(佛手柑)对急性焦虑无效 [B200].
方法论异质性
研究在石油选择、剂量、给药途径(吸入、按摩、局部)、暴露时间、结果测量和人群方面各不相同 [B323]. 这种异质性阻碍了综合和可比性 [B324].
缺少生物标志物
没有研究根据神经生物学亚型(例如神经炎症特征、E/I失衡标志物)对患者进行分层,或使用客观的生物标志物作为结果。 [B325]. 精准医疗方法可以识别响应者 [B326].
发表偏倚
正面案例报告和小规模试点研究比阴性研究更容易发表,这可能导致对效应的过度估计 [B327].
翻译差距
临床前模型(VPA大鼠、MIA小鼠)无法复制人类ASD异质性的全部复杂性 [B328]. 在动物研究中的萜烯剂量通常高于人体通过吸入暴露所能达到的水平 [B329].
安慰剂效应
芳香疗法与强烈的期望效应相关;由于明显的气味,盲法难以实现 [B330]. Bergamot-RCT 使用水作为安慰剂,这意味着对研究设盲不足。 [B200].
安全数据 关于儿童慢性吸入暴露的长期安全数据缺失 [B331]. 大多数安全数据来自局部或口服应用 [B332].
展望
研究重点
- 高质量的随机对照试验: 随机、双盲、安慰剂对照、具有足够效力(n > 100)、标准化油剂(GC/MS 认证)、客观结果指标(生物标志物、客观活动描记法、眼动追踪)和较长干预时间(8-12 周)的研究 [B333].
- 按亚型分层: 基于神经生物学亚型(神经炎症标志物、E/I失衡、微生物组特征)的响应者识别 [B334]. 精准医疗方法可能增加效应量 [B335].
- 机理研究 在干预前后进行包含生物标志物结果(细胞因子谱、神经影像、脑电图、微生物组分析)的人体研究,以验证机制 [B336].
- 药代动力学 吸入性萜烯暴露的人体药代动力学研究:血浆浓度、中枢神经系统渗透、代谢、剂量-效应关系 [B337].
- 联合疗法: 探究精油与既定干预措施(行为疗法、药物疗法、微生物组干预)之间的协同作用 [B338].
- 长期安全性 儿童慢性吸入性暴露的长期安全性前瞻性队列研究 [B339].
- 个性化芳香疗法: 基于个体症状特征、偏好和神经生物学标记的油品选取算法开发 [B340].
临床影响
基于现有证据,精油可作为多模式治疗方法的一部分,用于治疗自闭症谱系障碍 (ASD) 的共病症状(焦虑、睡眠障碍、感觉失调),但不应作为单一疗法或替代循证核心干预措施。 [B341]. 临床医生应:
- 沟通现实的期望:精油可以缓解合并症,但不能治愈核心症状 [B342]
- 考虑个人容忍度和偏好 [B343]
- 推荐优质的 GC/MS 认证油 [B344]
- 注意安全问题(癫痫、年龄、药物相互作用) [B345]
- 监测效果并调整治疗 [B346]
结论
精油是一种机制上合理但临床验证不足的自闭症谱系障碍辅助治疗选择。ASD 的神经生物学异质性——以 GABA/谷氨酸 E/I 不平衡、神经炎症、氧化应激、肠脑轴改变和突触功能障碍为特征——提供了多种治疗靶点,特种萜烯可以针对这些靶点。.
临床前证据表明,芳樟醇(GABA-A调节、血清素能调节)、β-石竹烯(CB2介导的神经炎症减轻)、乳香酸(Nrf2/HO-1激活)、1,8-桉叶素(抗焦虑GABA能作用)和依兰依兰(血清素/多巴胺调节)可以调节与ASD相关的病理生理机制,并在动物模型中改善ASD样行为。这种机制上的合理性得到了传统应用以及有限的人类焦虑、睡眠和情绪数据的支持。.
然而,ASD的临床证据质量较低:一项阴性随机对照试验(佛手柑),规模小但有方法学局限性的试点研究(薰衣草在ABA依从性方面)以及观察性研究。没有研究能够以足够的证据质量证明其对ASD核心症状的有效性。缺乏高质量、随机对照、使用标准化制剂、客观结局测量且具有足够统计效力的研究。.
为 Organik Aromas 3.0 扩香器开发了针对不同房间大小的剂量建议(2-10 滴用于 10-60 平方米,15-30 分钟,休息 30-60 分钟),并有适应不同时间的方案(早晨激活,晚上平静)以及针对 ASD 的特殊性(逐步引入,偏好筛选,感官过敏)。安全方面包括癫痫禁忌症(含樟脑的精油)、3 岁以下儿童禁忌(桉树精油)、与抗精神病药物的潜在相互作用(含佛手柑内酯的精油)以及需要 GC/MS 认证的纯精油。.
精油可作为多模式治疗方法的一部分,用于治疗自闭症谱系障碍(ASD)的共病症状(焦虑、睡眠障碍、感觉失调),但不能替代循证的核心干预措施(行为疗法、结构化教育)。未来的研究应优先考虑高质量的随机对照试验、基于神经生物学亚型的分层、机制性生物标志物研究和长期安全性数据,以验证精油在 ASD 治疗中的潜力并制定精准医疗方法。.
共同结论
跨领域见解:ADHD 与自闭症
注意缺陷多动障碍 (ADHD) 和自闭症谱系障碍 (ASD) 这两种神经发育障碍在基本神经生物学上存在共同点,这使得使用精油作为这两种疾病的辅助疗法具有合理性。.
共同的神经生物学目标结构
| 机制 | ADHD 相关性 | ASD 相关性 | 合适的油 |
| 多巴胺失调 | DAT过表达,多巴胺功能减退 | 中脑边缘系统功能障碍 | 迷迭香(1,8-桉树脑)、黑胡椒(β-石竹烯)、姜 |
| GABA/谷氨酸失衡 | 抑制性缺陷 | E/I-失衡(核心病理) | 薰衣草(芳樟醇)、香蜂草(迷迭香酸)、1,8-桉树脑、桉树 |
| 神经炎症 | 小胶质细胞激活 | TNF-α、IL-6、IL-1β升高 | β-石竹烯 (CB2),乳香 (Nrf2),生姜 (STAT3/TNF-α) |
| 氧化应激 | 线粒体功能障碍 | 谷胱甘肽降低,活性氧升高 | 乳香、薰衣草、生姜 |
| 血清素调节 | 去甲肾上腺素/血清素相互作用 | 5-HT失调 | 薰衣草、佛手柑、檀香 |
| Aufmerksamkeit/Kognition | 核心赤字(持续注意力) | 执行功能受影响 | 薄荷 (薄荷醇)、迷迭香、柠檬香蜂草、桉树 |
| 胆碱能通路 | 前额叶皮层 | 认知,学习 | 美兰nil (乙酰胆碱酯酶抑制剂),桉树 (1,8-桉叶油精) |
| 社会行为 | 较少影响 | 核心赤字 | 萜烯混合物(BTBR 模型),生姜(VPA 模型) |
通用应用建议(Organic Aromas 3.0 雾化器):
对于这两种适应症,无水无热的文丘里冷喷雾原理被认为是最佳的施用形式:– 萜烯无热降解 – 精确的颗粒大小,用于嗅觉吸收(< 5 µm) – GC/MS 纯净的精油保持其分子完整性
重要提示
精油辅助疗法不能替代循证医学标准疗法(ADHD 的哌甲酯、ASD 的 ABA/行为疗法)。它是一种补充性疗法,应个体化并经过专业指导。这两种适应症的证据尚有限;需要更大规模的随机对照试验。.
ADHD 参考
陈等人 (2008)。肉桂香薰疗法对注意力缺陷多动障碍儿童影响的研究。.https://pmc.ncbi.nlm.nih.gov/articles/PMC8538900/
陈等人。肉桂香薰疗法对注意缺陷多动障碍儿童的影响。.https://pmc.ncbi.nlm.nih.gov/articles/PMC8538900/
[A3] Godfrey (2001)。精油在注意力缺陷多动障碍的治疗和管理中的作用。《国际芳香疗法杂志》。.https://doi.org/10.1016/S0962-4562(01 )80035-7
[A4] Godfrey. 精油:注意力缺陷多动障碍(ADHD)的辅助治疗。.https://www.researchgate.net/publication/248871372_The_role_of_essential_oils_in_the_treatment_and_management_of_attention_deficit_hyperactive_disorder
[A5] Godfrey. 精油在注意力缺陷多动障碍(ADHD)的治疗和管理中的作用。.https://www.researchgate.net/publication/248871372_The_role_of_essential_oils_in_the_treatment_and_management_of_attention_deficit_hyperactive_disorder
[A6] 丁等人(1994)。[LLC]dl-苏糖甲基苯丙胺类对突触前多巴胺能神经元的药代动力学和体内特异性。Synapse。.https://doi.org/10.1002/SYN.890180207
[A7] Volkow 等人 (2002)。哌甲酯的作用机制:正电子发射断层扫描研究的见解。注意缺陷杂志。.https://doi.org/10.1177/070674370200601S05
[A8] Silvа (2024)。芳香疗法在患有注意力缺陷多动障碍和特应性皮炎的非典型儿童护理中的应用。.https://doi.org/10.62435/2965-7253.bjhae.2024.iisinaroma1
[A9] Volkow等 (1998)。口服哌甲酯治疗剂量诱导的人类大脑多巴胺转运体占有率。American Journal of Psychiatry。.https://doi.org/10.1176/AJP.155.10.1325
[A10] Jacob 等人(2024)。芳香疗法在学生支持中的应用。.https://doi.org/10.37885/240717094
[A11] Dipasquale 等人 (2020)。揭示哌甲酯对多巴胺能和去甲肾上腺素能功能回路的影响。《生物档案》预印本。.https://doi.org/10.1101/2020.03.09.983528
[A12] Volkow 等人 (2002)。多巴胺在人类甲基苯丙胺的治疗和强化作用中的作用:来自影像学研究的结果。《欧洲神经精神药理学》。.https://doi.org/10.1016/S0924-977X(02 )00104-9
[A13] Spencer 等人 (2006)。一项 PET 研究,考察短期和长期口服哌甲酯的药代动力学、检测和喜好度以及多巴胺转运体受体占据。American Journal of Psychiatry。.https://doi.org/10.1176/APPI.AJP.163.3.387
[A14] Castillo 等人。(2018)。精油作为人类行为的调节剂。《热带亚热带农田生态系统》。.https://www.revista.ccba.uady.mx/ojs/index.php/TSA/article/view/2583
[A15] Crunelle et al. (2014)。哌甲酯多巴胺转运体占有率和成人ADHD的冲动性。《英国精神病学杂志》。.https://doi.org/10.1192/BJP.BP.113.132977
[A16] 丁等(1997)。手性药物:[¹¹C]右旋苏-和左旋苏-哌甲酯在人脑和狒狒脑中药代动力学比较。《精神药理学》。.https://doi.org/10.1007/S002130050267
[A17] Spencer 等人 (2009)。一项 PET 研究,考察两种长效哌甲酯制剂在成人中的药代动力学和多巴胺转运体占有率。《国际分子医学杂志》。.https://doi.org/10.3892/IJMM_00000339
[A18] Marcel 等人 (2024)。纹状体多巴胺系统对哌甲酯的反应性——一项在患有多动症的男性儿童和青少年中的受试者内 I-123-β-CIT-SPECT 研究。.https://doi.org/10.5283/epub.57629
[A19] Kennedy et al. (2018)。挥发性萜烯与大脑功能:薄荷精油对认知和情绪影响的探究及其体外性质与中枢神经系统功能的相关性。《营养素》。.https://doi.org/10.3390/NU10081029
[A20] Batiha 等人 (2023)。薰衣草物种生物活性成分和药理特性的综述。Naunyn-Schmiedeberg's Archives of Pharmacology。.https://doi.org/10.1007/s00210-023-02392-x
[A21] Guzmán-Gutiérrez 等人 (2015)。芳樟醇和 β-蒎烯通过单胺能通路发挥其抗抑郁样活性。.https://pubmed.ncbi.nlm.nih.gov/25771248/
[A22] Martinez 等人 (2020)。腹侧纹状体支持哌甲酯在时间折扣任务中表现出的冲动选择的治疗效果。《科学报告》。.https://doi.org/10.1038/s41598-020-57595-6
[A23] López 等人。(2017)。探索薰衣草 (Lavandula angustifolia) 精油对中枢神经系统靶点的药理机制。Frontiers in Pharmacology。.https://doi.org/10.3389/FPHAR.2017.00280
[A24] Rathnayake (2025) 。“神经芳香疗法”:芳香疗法与边缘系统应激反应。.https://doi.org/10.4038/jssnisd.v3i1.28
[A25] Aster 等人(2021)。纹状体多巴胺系统对哌甲酯的反应性——一项在注意力缺陷/多动障碍儿童和青少年中进行的 I-123-β-CIT-SPECT 单个受试者研究。《medRxiv》。.https://doi.org/10.1101/2021.11.19.21265993
[A26] Cardoso 等人(2025)。薰衣草精油对人类神经系统的影响:一项综述。《已删除期刊》。.https://doi.org/10.62435/2965-7253.bjhae.2025.bjhae22
[A27] Moss 等人 (2003)。迷迭香和薰衣草精油的香气对健康成年人的认知和情绪有不同的影响。《神经科学国际杂志》。.https://doi.org/10.1080/00207450390161903
[A28] Sinha 等人 (2005)。儿童注意力缺陷多动障碍的补充与替代医学使用。《儿科与儿童健康杂志》。.https://doi.org/10.1111/J.1440-1754.2005.00530.X
[A29] Springer 等人 (2018)。ADHD:生命周期及其从青少年到成年期的影响。.https://dergipark.org.tr/en/pub/pgy/548052.xml
[A30] Colzato等人 (2014)。气味调节注意力闪烁中的注意力控制。《注意、知觉与心理物理学》.https://doi.org/10.3758/S13414-014-0733-0
A31. Kennedy, D. O., Scholey, A., Tildesley, N. T. J., Perry, E. K., & Wesnes, K. A. (2002). 镇静安神,提高认知能力——急性服用香蜂草(柠檬香膏)后的情绪和认知表现的调节。. 药理学、生物化学与行为学, 72(4), 953–964. https://doi.org/10.1016/S0091-3057(02)00777-3
A32. Müller, S. F., & Klement, S. (2006). 缬草和柠檬香蜂草的组合在治疗儿童烦躁不安和睡眠困难方面有效。. 植物药理学, 13(6), 383–387. https://doi.org/10.1016/j.phymed.2006.01.013
A33. Scholey, A., Gibbs, A., Neale, C., et al. (2014). 柠檬香蜂草食品的抗应激作用。. 营养素, 6(11), 4805–4821. https://doi.org/10.3390/nu6114805
A34. 桉树和茉莉精油对认知功能和阿尔茨海默病相关病理的神经保护作用的研究综述。(2022)。. 神经保护性精油综述.
参考 ASD
[B1] Maenner MJ, Shaw KA, Bakian AV,等人。 8岁儿童自闭症谱系障碍的患病率和特征——自闭症和发育障碍监测网络,美国11个地点,2018年。MMWR Surveill Summ。2021;70(11):1-16。.https://doi.org/10.15585/mmwr.ss7011a1
[B2] Zeidan J, Fombonne E, Scorah J, 等。自闭症的全球患病率:系统评价更新。Autism Res. 2022;15(5):778-790。.https://doi.org/10.1002/aur.2696
[B3] Lord C, Elsabbagh M, Baird G, Veenstra-Vanderweele J. 孤独症谱系障碍. 柳叶刀. 2018;392(10146):508-520.https://doi.org/10.1016/S0140-6736(18 )31129-2
[B4] Masi A, DeMayo MM, Glozier N, Guastella AJ. 自闭症谱系障碍、异质性及治疗选择概述。神经科学通讯。2017;33(2):183-193。.https://doi.org/10.1007/s12264-017-0100-y
[B5] 美国精神病学协会. 精神疾病诊断与统计手册,第五版 (DSM-5). 阿灵顿,弗吉尼亚州:美国精神病学出版社;2013。.https://www.psychiatry.org/psychiatrists/practice/dsm
[B6] Marco EJ, Hinkley LB, Hill SS, Nagarajan SS。自闭症中的感觉处理:神经生理学研究回顾。儿科研究。2011;69(5 Pt 2):48R-54R。.https://doi.org/10.1203/PDR.0b013e3182130c54
[B7] Baum SH, Stevenson RA, Wallace MT. 自闭症谱系障碍中的感觉和多感觉功能的行为、感知和神经改变。Prog Neurobiol. 2015;134:140-160。.https://doi.org/10.1016/j.pneurobio.2015.09.007
[B8] van Steensel FJ, Bögels SM, Perrin S. 儿童和青少年自闭症谱系障碍的焦虑障碍:一项荟萃分析。临床儿童家庭心理学评论。2011;14(3):302-317。.https://doi.org/10.1007/s10567-011-0097-0
[B9] Souders MC, Mason TB, Valladares O, 等人。自闭谱系障碍儿童的睡眠行为和睡眠质量。睡眠。2009;32 (12):1566-1578。.https://doi.org/10.1093/sleep/32.12.1566
[B10] McElhanon BO, McCracken C, Karpen S, Sharp WG. 孤独症谱系障碍的胃肠道症状:一项荟萃分析。《儿科学》。2014;133(5):872-883。.https://doi.org/10.1542/peds.2013-3995
[B11] Geschwind DH, State MW. 孤独症谱系障碍中的基因搜寻:迈向精准医疗之路。《柳叶刀·神经病学》,2015;14(11):1109-1120.https://doi.org/10.1016/S1474-4422(15 )00044-7
[B12] De Rubeis S, He X, Goldberg AP, 等人. 哪些突触、转录和染色质基因在自闭症中被破坏. [Nature]. 2014;515(7526):209-215.https://doi.org/10.1038/nature13772
[B13] Sandin S, Lichtenstein P, Kuja-Halkola R, 等. 自闭症谱系障碍的遗传力. JAMA. 2017;318(12):1182-1184.https://doi.org/10.1001/jama.2017.12141
[B14] Rubenstein JL, Merzenich MM. 自闭症模型:关键神经系统中兴奋/抑制比例增加。《基因、大脑与行为》(Genes Brain Behav)。2003;2(5):255-267。.https://doi.org/10.1034/j.1601-183x.2003.00037.x
[B15] Vargas DL, Nascimbene C, Krishnan C, Zimmerman AW, Pardo CA. 神经胶质细胞激活与自闭症患者大脑中的神经炎症。Ann Neurol. 2005;57(1):67-81。.https://doi.org/10.1002/ana.20315
[B16] Rossignol DA, Frye RE. 证据表明氧化应激、线粒体功能障碍和炎症在大脑自闭症患者中的联系。Front Physiol. 2014;5:150.https://doi.org/10.3389/fphys.2014.00150
[B17] Nelson SB, Valakh V. 兴奋/抑制平衡与自闭症谱系障碍中的电路稳态. Neuron. 2015;87(4):684-698.https://doi.org/10.1016/j.neuron.2015.07.033
[B18] Lee E, Lee J, Kim E. 孤独症谱系障碍动物模型的兴奋/抑制失衡。Biol Psychiatry。2017;81(10):838-847。.https://doi.org/10.1016/j.biopsych.2016.05.011
[B19] Horder J, Lavender T, Mendez MA, et al. 成人自闭症谱系障碍中皮层下谷氨酸/谷氨酰胺水平降低:一项 [¹H]MRS 研究。Transl Psychiatry。2013;3:e279。.https://doi.org/10.1038/tp.2013.53
[B20] Gaetz W, Bloy L, Wang DJ, 等人。自闭症谱系儿童大脑中的GABA估计:测量精度和区域皮层差异。Neuroimage。2014;86:1-9。.https://doi.org/10.1016/j.neuroimage.2013.05.068
[B21] Rojas DC, Singel D, Steinmetz S, Hepburn S, Brown MS. 自闭症儿童及其未患病同胞的左侧外侧裂GABA浓度降低。《神经影像学》. 2014;86:28-34.https://doi.org/10.1016/j.neuroimage.2013.01.045
[B22] Amaral DG, Schumann CM, Nordahl CW. 神经解剖学与自闭症. Trends Neurosci. 2008;31(3):137-145.https://doi.org/10.1016/j.tins.2007.12.005
[B23] Courchesne E, Pierce K. 为什么自闭症中的额叶皮层可能只在“自言自语”:局部过度连接但长距离断开。Curr Opin Neurobiol. 2005;15(2):225-230。.https://doi.org/10.1016/j.conb.2005.03.001
[B24] Baron-Cohen S, Ring HA, Bullmore ET, Wheelwright S, Ashwin C, Williams SC. 自闭症的杏仁核理论. 神经科学与行为评论. 2000;24(3):355-364.https://doi.org/10.1016/s0149-7634(00 )00011-7
[B25] Estes ML, McAllister AK. 母体免疫激活:对神经精神疾病的影响。《科学》。2016;353(6301):772-777。.https://doi.org/10.1126/science.aag3194
[B26] Matta SM, Hill-Yardin EL, Crack PJ. 神经炎症对自闭症谱系障碍的影响。Brain Behav Immun。2019;79:75-90。.https://doi.org/10.1016/j.bbi.2019.04.037
[B27] Careaga M, Murai T, Bauman MD. 母体免疫激活与自闭症谱系障碍:从啮齿动物到非人及人类灵长类动物. Biol Psychiatry. 2017;81(5):391-401.https://doi.org/10.1016/j.biopsych.2016.10.020
[B28] Chauhan A, Chauhan V. 氧化应激在自闭症中的作用. 病理生理学. 2006;13(3):171-181.https://doi.org/10.1016/j.pathophys.2006.05.007
[B29] Rose S, Melnyk S, Pavliv O, et al. 氧化损伤和炎症与自闭症大脑中谷胱甘肽氧化还原状态低有关。Transl Psychiatry. 2012;2:e134。.https://doi.org/10.1038/tp.2012.61
[B30] Kang DW, Adams JB, Gregory AC, et al. 微生态菌群移植疗法改变肠道生态系统并改善胃肠道和自闭症症状:一项开放标签研究。《微生物组》。2017;5(1):10。.https://doi.org/10.1186/s40168-016-0225-7
[B31] Vuong HE, Hsiao EY. 肠道微生物组在自闭症谱系障碍中的新兴作用。Biol Psychiatry。2017;81(5):411-423。.https://doi.org/10.1016/j.biopsych.2016.08.024
[B32] Sharon G, Cruz NJ, Kang DW, 等. 来自自闭症谱系障碍的人类肠道微生物群在小鼠中促成行为症状。Cell。2019;177(6):1600-1618.e17。.https://doi.org/10.1016/j.cell.2019.05.004
[B33] Sgritta M, Dooling SW, Buffington SA, 等人。自闭症谱系障碍小鼠模型中微生物介导的社会行为改变的潜在机制。Neuron。2019;101(2):246-259.e6。.https://doi.org/10.1016/j.neuron.2018.11.018
[B34] Reichow B, Hume K, Barton EE, Boyd BA. 针对自闭症谱系障碍 (ASD) 幼儿的早期强化行为干预 (EIBI)。Cochrane Database Syst Rev. 2018;5(5):CD009260。.https://doi.org/10.1002/14651858.CD009260.pub3
[B35] Dawson G, Rogers S, Munson J, 等人。一项针对自闭症幼儿干预试验的随机对照试验:早期人际交流模型。儿科学。2010;125(1):e17-23。.https://doi.org/10.1542/peds.2009-0958
[B36] McCracken JT, McGough J, Shah B, et al. 瑞斯匹瑞酮治疗有严重行为问题的自闭症儿童。 N Engl J Med. 2002;347(5):314-321。.https://doi.org/10.1056/NEJMoa013171
[B37] Marcus RN,Owen R,Kamen L 等人。阿立哌唑治疗自闭症相关易怒儿童和青少年的一项安慰剂对照、固定剂量研究。J Am Acad Child Adolesc Psychiatry。2009;48(11):1110-1119。.https://doi.org/10.1097/CHI.0b013e3181b76658
[B38] Scahill L, McDougle CJ, Aman MG, 等人。利培酮和父母培训对广泛性发育障碍和严重行为问题儿童适应性功能的影响。《美国儿童青少年精神病学院学报》2012;51(2):136-146。.https://doi.org/10.1016/j.jaac.2011.11.010
[B39] Rossignol DA, Frye RE. 褪黑激素在自闭症谱系障碍中的应用:系统性回顾与荟萃分析. Dev Med Child Neurol. 2011;53(9):783-792.https://doi.org/10.1111/j.1469-8749.2011.03980.x
[B40] Hollander E, Soorya L, Chaplin W, et al. 一项关于氟西汀治疗成年自闭症谱系障碍重复行为和整体严重程度的双盲安慰剂对照试验。Am J Psychiatry。2012;169(3):292-299。.https://doi.org/10.1176/appi.ajp.2011.10050764
[B41] Spencer D, Marshall J, Post B, et al. 儿童自闭症谱系障碍的神经精神药物使用及多重用药情况。《儿科学》. 2013;132(5):833-840.https://doi.org/10.1542/peds.2012-3774
[B42] Koulivand PH, Khaleghi Ghadiri M, Gorji A. 薰衣草与神经系统. 循证补充替代医学. 2013;2013:681304.https://doi.org/10.1155/2013/681304
[B43] Sowndhararajan K, Kim S. 香料对人类精神生理活动的影响:以人类脑电图反应为例。Sci Pharm. 2016;84(4):724-751。.https://doi.org/10.3390/scipharm84040724
[B44] Ali B, Al-Wabel NA, Shams S, Ahamad A, Khan SA, Anwar F. 用于芳香疗法的精油:系统性评价。亚洲太平洋热带生物医学杂志。2015;5(8):601-611。.https://doi.org/10.1016/j.apjtb.2015.05.007
[B45] Sohal VS, Rubenstein JLR. 兴奋-抑制平衡作为研究神经精神障碍机制的框架。《精神病学分子》. 2019;24(9):1248-1257.https://doi.org/10.1038/s41380-019-0426-0
[B46] Antoine MW, Langberg T, Schnepel P, Feldman DE。兴奋-抑制比增加可稳定四种孤独症小鼠模型的突触和电路兴奋性,《神经元》。2019;101(4):648-661.e4。.https://doi.org/10.1016/j.neuron.2018.12.026
[B47] Fatemi SH, Halt AR, Stary JM, Kanodia R, Schulz SC, Realmuto GR. 谷氨酸脱羧酶65和67 kDa蛋白在自闭症顶叶和大脑皮层中减少。Biol Psychiatry. 2002;52(8):805-810。.https://doi.org/10.1016/s0006-3223(02 )01430-0
[B48] Yip J, Soghomonian JJ, Blatt GJ. 焦虑症小脑浦肯野细胞中GAD67 mRNA水平降低:病理生理学意义。Acta Neuropathol. 2007;113(5):559-568。.https://doi.org/10.1007/s00401-006-0176-3
[B49] Ben-Ari Y. 兴奋性GABA在发育中的作用:后天环境的本质。Nat Rev Neurosci. 2002;3(9):728-739。.https://doi.org/10.1038/nrn920
[B50] Tyzio R, Nardou R, Ferrari DC, 等. 分娩过程中催产素介导的GABA抑制可减轻啮齿动物后代的自闭症发病. 科学. 2014;343(6171):675-679.https://doi.org/10.1126/science.1247190
[B51] Lemonnier E, Degrez C, Phelep M, et al. A randomised controlled trial of bumetanide in the treatment of autism in children. Transl Psychiatry. 2012;2:e202.https://doi.org/10.1038/tp.2012.124
[B52] Lemonnier E, Villeneuve N, Sonie S, et al. 丁丁酰肼对自闭症谱系障碍儿童和青少年神经行为功能的影响。Transl Psychiatry。2017;7(3):e1056。.https://doi.org/10.1038/tp.2017.10
[B53] Carlson GC. 谷氨酸受体功能障碍和自闭症谱系障碍模型中的药物靶点. 药理学行为学报. 2012;100(4):850-854.https://doi.org/10.1016/j.pbb.2011.02.003
[B54] Uzunova G, Pallanti S, Hollander E. 孤独症谱系障碍中的兴奋/抑制失衡:对干预和治疗的启示. World J Biol Psychiatry. 2016;17(3):174-186.https://doi.org/10.3109/15622975.2015.1085597
[B55] Yoo HJ, Cho IH, Park M, Yang SY, Kim SA. 全家为基础的 GRIN2A 和 GRIN2B 与韩国自闭症谱系障碍的关联。Neurosci Lett。2012;512(2):89-93。.https://doi.org/10.1016/j.neulet.2012.01.061
[B56] Silverman JL, Tolu SS, Barkan CL, Crawley JN. BTBR小鼠自闭症模型中的重复性自我梳理行为被mGluR5拮抗剂MPEP阻断。Neuropsychopharmacology。2010;35(4):976-989。.https://doi.org/10.1038/npp.2009.201
[B57] Muller CL, Anacker AMJ, Veenstra-VanderWeele J. 自闭症谱系障碍中的血清素系统:从生物标志物到动物模型. Neuroscience. 2016;321:24-41.https://doi.org/10.1016/j.neuroscience.2015.11.010
[B58] Pavăl D. A Dopamine Hypothesis of Autism Spectrum Disorder. Dev Neurosci. 2017;39(5):355-360.https://doi.org/10.1159/000478725
[B59] Schain RJ, Freedman DX. Studies on 5-hydroxyindole metabolism in autistic and other mentally retarded children. J Pediatr. 1961;58:315-320.https://doi.org/10.1016/s0022-3476(61 )80261-8
[B60] Cook EH Jr, Leventhal BL. 自闭症中的血清素系统. Curr Opin Pediatr. 1996;8(4):348-354.https://doi.org/10.1097/00008480-199608000-00008
[B61] Chugani DC, Muzik O, Behen M,等. 自闭症儿童和非自闭症儿童大脑血清素合成能力的发育变化. Ann Neurol. 1999;45(3):287-295.https://doi.org/10.1002/1531-8249(199903 )45:33.0.co;2-9
[B62] Devlin B, Cook EH Jr, Coon H, 等. 自闭症与5-羟色胺转运蛋白:漫长而短暂的故事. Mol Psychiatry. 2005;10(12):1110-1116.https://doi.org/10.1038/sj.mp.4001724
[B63] Whitaker-Azmitia PM. 行为和细胞学后果,在脑发育期间增加血清素能活性:在自闭症中起作用吗? Int J Dev Neurosci. 2005;23(1):75-83.https://doi.org/10.1016/j.ijdevneu.2004.07.022
[B64] King BH, Hollander E, Sikich L, 等. 西酞普兰在患有自闭症谱系障碍和高度重复行为的儿童中无效:西酞普兰对自闭症儿童无效。《精神病学年鉴》。2009;66(6):583-590。.https://doi.org/10.1001/archgenpsychiatry.2009.30
[B65] Dichter GS, Damiano CA, Allen JA。精神和神经发育障碍和遗传综合征中的奖赏回路功能障碍:动物模型和临床发现。J Neurodev Disord。2012;4(1):19。.https://doi.org/10.1186/1866-1955-4-19
[B66] Kohls G, Schulte-Rüther M, Nehrkorn B, et al. 孤独症谱系障碍中的奖赏系统功能障碍。Soc Cogn Affect Neurosci。2013;8(5):565-572。.https://doi.org/10.1093/scan/nss033
[B67] Delmonte S, Balsters JH, McGrath J, 等人。孤独症谱系障碍中的社会和金钱奖励处理。Mol Autism。2012;3(1):7。.https://doi.org/10.1186/2040-2392-3-7
[B68] 中村A,关根Y,大内Y等人。高功能自闭症成人大脑中的血清素和多巴胺转运体结合。精神病学年鉴。2010;67(1):59-68。.https://doi.org/10.1001/archgenpsychiatry.2009.137
[B69] Staal WG. 自闭症、DRD3与重复刻板行为:当前知识概述。Eur Neuropsychopharmacol。2015;25(9):1421-1426。.https://doi.org/10.1016/j.euroneuro.2014.08.011
[B70] Gunaydin LA, Grosenick L, Finkelstein JC, 等人。揭示社交行为的自然神经投射动力学。Cell。2014;157(7):1535-1551。.https://doi.org/10.1016/j.cell.2014.05.017
[B71] Insel TR. 神经科学中翻译的挑战:催产素、加压素和亲和行为的元分析。《神经元》杂志。2010;65(6):768-779。.https://doi.org/10.1016/j.neuron.2010.03.005
[B72] Modahl C, Green L, Fein D, 等. 自闭症儿童血浆催产素水平. Biol Psychiatry. 1998;43(4):270-277.https://doi.org/10.1016/s0006-3223(97 )00439-3
[B73] Gregory SG, Connelly JJ, Towers AJ, et al. 催产素受体缺乏在自闭症中的基因组和表观遗传学证据。BMC Med. 2009;7:62。.https://doi.org/10.1186/1741-7015-7-62
[B74] Guastella AJ, Einfeld SL, Gray KM 等人。鼻腔催产素可改善自闭症谱系障碍青少年的情绪识别。Biol Psychiatry。2010;67(7):692-694。.https://doi.org/10.1016/j.biopsych.2009.09.020
[B75] Parker KJ, Oztan O, Libove RA, 等人。鼻腔催产素治疗自闭症儿童的社交缺陷和反应生物标志物。《美国国家科学院院刊》2017;114(30):8119-8124。.https://doi.org/10.1073/pnas.1705521114
[B76] Peñagarikano O, Lázaro MT, Lu XH, 等. 外源性和诱发催产素可恢复自闭症Cntnap2小鼠模型的社会行为。Sci Transl Med。2015;7(271):271ra8。.https://doi.org/10.1126/scitranslmed.3010257
[B77] Pardo CA, Vargas DL, Zimmerman AW. 免疫、神经胶质细胞和自闭症中的神经炎症。Int Rev Psychiatry。2005;17(6):485-495。.https://doi.org/10.1080/02646830500381930
[B78] Edmonson C, Ziats MN, Rennert OM. 自闭症死后前额叶皮层和小脑中胶质细胞标志物表达的改变。Mol Autism。2014;5:3。.https://doi.org/10.1186/2040-2392-5-3
[B79] Salter MW, Stevens B. 小胶质细胞成为脑部疾病的关键参与者. Nat Med. 2017;23(9):1018-1027.https://doi.org/10.1038/nm.4397
[B80] Morgan JT, Chana G, Pardo CA, et al. 孤独症背外侧前额叶皮层观察到的小胶质细胞激活和密度增加。Biol Psychiatry. 2010;68(4):368-376.https://doi.org/10.1016/j.biopsych.2010.05.024
[B81] Suzuki K, Sugihara G, Ouchi Y, 等. 谱系障碍年轻成人中的小胶质细胞激活. JAMA Psychiatry. 2013;70(1):49-58.https://doi.org/10.1001/jamapsychiatry.2013.272
[B82] Zürcher NR, Loggia ML, Lawson R, 等。 经 [(11)C]-PBR28 评估,肌萎缩侧索硬化症患者体内胶质细胞激活增加。 Neuroimage Clin. 2015;7:409-414。.https://doi.org/10.1016/j.nicl.2015.01.009
[B83] Schafer DP, Lehrman EK, Kautzman AG, et al. 小胶质细胞以激活和补体依赖的方式雕塑出生后神经回路。Neuron。2012;74(4):691-705。.https://doi.org/10.1016/j.neuron.2012.03.026
[B84] Ashwood P, Krakowiak P, Hertz-Picciotto I, Hansen R, Pessah I, Van de Water J. 自闭症谱系障碍中升高的血浆细胞因子为免疫功能障碍提供了证据,并与行为结局受损有关。Brain Behav Immun。2011;25(1):40-45。.https://doi.org/10.1016/j.bbi.2010.08.003
[B85] Li X, Chauhan A, Sheikh AM, 等。孤独症患者大脑中免疫反应升高。J Neuroimmunol。2009;207(1-2):111-116。.https://doi.org/10.1016/j.jneuroim.2008.12.002
[B86] Smith SE, Li J, Garbett K, Mirnics K, Patterson PH. 母体免疫激活通过白介素6改变胎儿大脑发育. J Neurosci. 2007;27(40):10695-10702.https://doi.org/10.1523/JNEUROSCI.2178-07.2007
[B87] 崔GB, 任YS, 王H, 等. 小鼠母体白介素-17a通路促进后代自闭症样表型. 科学. 2016;351(6276):933-939.https://doi.org/10.1126/science.aad0314
[B88] Stellwagen D, Malenka RC. 神经突触缩放由胶质细胞 TNF-α 介导。自然。2006;440(7087):1054-1059。.https://doi.org/10.1038/nature04671
[B89] 魏 H、Chadman KK、McCloskey DP 等人。脑 IL-6 升高导致神经回路失衡并介导自闭症样行为。Biochem Biophys Acta. 2012;1822(6):831-842。.https://doi.org/10.1016/j.bbadis.2012.01.011
[B90] Verkhratsky A,Nedergaard M。星形胶质细胞的生理学。Physiol Rev。2018;98(1):239-389。.https://doi.org/10.1152/physrev.00042.2016
[B91] Laurence JA, Fatemi SH. 自闭症患者的额上回、顶叶和脑叶皮层中胶质纤维酸性蛋白升高。小脑。2005;4(3):206-210。.https://doi.org/10.1080/14734220500208846
[B92] Rothstein JD, Dykes-Hoberg M, Pardo CA, 等. 谷氨酸转运蛋白的基因敲除揭示了星形胶质细胞转运在兴奋性毒性和谷氨酸清除中的重要作用. Neuron. 1996;16(3):675-686.https://doi.org/10.1016/s0896-6273(00 )80086-0
[B93] Sies H, Jones DP. 活性氧(ROS)作为多效生理信号分子。Nat Rev Mol Cell Biol。2020;21(7):363-383。.https://doi.org/10.1038/s41580-020-0230-3
Frustaci A, Neri M, Cesario A, 等人。自闭症中与氧化应激相关的生物标志物:系统评价和荟萃分析。自由基生物医学杂志。2012;52(10):2128-2141。.https://doi.org/10.1016/j.freeradbiomed.2012.03.011
[B95] Bjørklund G, Meguid NA, El-Bana MA, 等人。自闭症谱系障碍中的氧化应激。Mol Neurobiol。2020;57(5):2314-2332。.https://doi.org/10.1007/s12035-019-01742-2
[B96] James SJ, Cutler P, Melnyk S, 等. 自闭症儿童氧化应激增加和甲基化能力受损的代谢生物标志物. Am J Clin Nutr. 2004;80(6):1611-1617.https://doi.org/10.1093/ajcn/80.6.1611
[B97] Rose S, Melnyk S, Pavliv O, et al. 氧化损伤和炎症与自闭症大脑中谷胱甘肽氧化还原状态低的关联证据。Transl Psychiatry. 2012;2:e134.https://doi.org/10.1038/tp.2012.61
[B98] Bowers K, Li Q, Bressler J, 等人. 谷胱甘肽通路基因变异与自闭症谱系障碍的风险. J Neurodev Disord. 2011;3(2):132-143.https://doi.org/10.1007/s11689-011-9077-4
[B99] Rossignol DA, Frye RE. 孤独症谱系障碍中的线粒体功能障碍:系统回顾与荟萃分析. Mol Psychiatry. 2012;17(3):290-314.https://doi.org/10.1038/mp.2010.136
[B100] Giulivi C, Zhang YF, Omanska-Klusek A, et al. 细胞线粒体功能障碍在自闭症中的作用. JAMA. 2010;304(21):2389-2396.https://doi.org/10.1001/jama.2010.1706
[B101] Duchen MR. 细胞信号传导到细胞死亡中的线粒体和钙。J Physiol。2000;529 Pt 1(Pt 1):57-68。.https://doi.org/10.1111/j.1469-7793.2000.00057.x
[B102] Bambini-Junior V, Rodrigues L, Behr GA,等人。孕期缬氨酸暴露诱导的自闭症动物模型:行为改变和肝脏参数。Brain Res. 2011;1408:8-16。.https://doi.org/10.1016/j.brainres.2011.06.015
[B103] Ma Q. Nrf2在氧化应激和毒性中的作用. Annu Rev Pharmacol Toxicol. 2013;53:401-426.https://doi.org/10.1146/annurev-pharmtox-011112-140320
[B104] Singh K, Connors SL, Macklin EA, et al. 萝卜硫磺治疗自闭症谱系障碍 (ASD)。Proc Natl Acad Sci U S A. 2014;111(43):15550-15555。.https://doi.org/10.1073/pnas.1416940111
[B105] Bent S, Lawton B, Warren T, 等. 确定与西兰花硫代葡萄糖苷治疗的自闭症儿童临床改善相关的尿液代谢物. Mol Autism. 2018;9:35.https://doi.org/10.1186/s13229-018-0218-4
[B106] Cryan JF, O’Riordan KJ, Cowan CSM, 等. 菌群-肠道-脑轴. 生理学评论. 2019;99(4):1877-2013.https://doi.org/10.1152/physrev.00018.2018
[B107] Chaidez V, Hansen RL, Hertz-Picciotto I. 自闭症、发育迟缓或正常发育儿童的胃肠道问题. J Autism Dev Disord. 2014;44(5):1117-1127.https://doi.org/10.1007/s10803-013-1973-x
[B108] Adams JB, Johansen LJ, Powell LD, Quig D, Rubin RA. 儿童肠道菌群与肠道状况——与普通儿童的比较及与自闭症严重程度的相关性。BMC Gastroenterol. 2011;11:22。.https://doi.org/10.1186/1471-230X-11-22
[B109] 许M,许X,李J,李F. 肠道微生物群与自闭症谱系障碍的关联:一项系统评价和荟萃分析。《精神病学前沿》。2019;10:473。.https://doi.org/10.3389/fpsyt.2019.00473
[B110] 刘 F, 李 J, 吴 F, 等. 自闭症谱系障碍中肠道微生物群组成的改变和功能:一项系统评价。Transl Psychiatry。2019;9(1):43。.https://doi.org/10.1038/s41398-019-0389-6
[B111] 康迪惠、朴政国、伊尔汗ZE等。自闭症儿童肠道菌群中普雷沃菌和其他发酵菌的发生率降低。《公共科学图书馆·综合》2013;8(7):e68322。.https://doi.org/10.1371/journal.pone.0068322
[B112] Stilling RM, van de Wouw M, Clarke G, Stanton C, Dinan TG, Cryan JF. 丁酸的神经药理学:微生物-肠道-大脑轴的“老本行”? Neurochem Int. 2016;99:110-132.https://doi.org/10.1016/j.neuint.2016.06.011
[B113] MacFabe DF, Cain DP, Rodriguez-Capote K, 等人。脑室内丙酸在大鼠中的神经生物学效应:短链脂肪酸在自闭症谱系障碍的发病机制和特征中的可能作用。行为脑研究。2007;176(1):149-169。.https://doi.org/10.1016/j.bbr.2006.07.025
[B114] 萧EY, McBride SW, Hsien S, 等. 菌群调节与神经发育障碍相关的行为和生理异常. Cell. 2013;155(7):1451-1463.https://doi.org/10.1016/j.cell.2013.11.024
[B115] Bonaz B, Bazin T, Pellissier S. 迷走神经在肠道菌群-肠道-大脑轴中的作用. Front Neurosci. 2018;12:49.https://doi.org/10.3389/fnins.2018.00049
[B116] 刘S、萩原SI、巴格瓦A。生命早期逆境、表观遗传学和内脏高敏性。神经胃肠病学与运动学。2017;29(9)。.https://doi.org/10.1111/nmo.13170
[B117] Zoghbi HY, Bear MF. 神经发育障碍伴自闭症和智力障碍的突触功能障碍。Cold Spring Harb Perspect Biol. 2012;4(3):a009886.https://doi.org/10.1101/cshperspect.a009886
[B118] Bourgeron T. 从遗传结构到自闭症谱系障碍的突触可塑性。Nat Rev Neurosci。2015;16(9):551-563。.https://doi.org/10.1038/nrn3992
Südhof TC. 神经粘蛋白和神经突触蛋白连接突触功能与认知疾病. Nature. 2008;455(7215):903-911.https://doi.org/10.1038/nature07456
[B120] Jamain S, Quach H, Betancur C, 等人。 编码神经纤维蛋白 NLGN3 和 NLGN4 的 X 连锁基因突变与自闭症相关。 Nat Genet. 2003;34(1):27-29。.https://doi.org/10.1038/ng1136
[B121] Tabuchi K, Blundell J, Etherton MR, 等人。一项与自闭症相关的神经细胞粘着分子3突变增加了小鼠的抑制性突触传递。科学。2007;318(5847):71-76。.https://doi.org/10.1126/science.1146221
[B122] Jamain S, Radyushkin K, Hammerschmidt K, 等人. 单基因遗传自闭症小鼠模型中社交互动减少和超声波交流障碍. Proc Natl Acad Sci U S A. 2008;105(5):1710-1715.https://doi.org/10.1073/pnas.0711555105
[B123] Grabrucker AM, Schmeisser MJ, Schoen M, Boeckers TM. 突触后 ProSAP/Shank 骨架在突触病变中的十字准线. Trends Cell Biol. 2011;21(10):594-603.https://doi.org/10.1016/j.tcb.2011.07.003
[B124] Phelan K, McDermid HE. 22q13.3缺失综合征(Phelan-McDermid综合征)。Mol Syndromol. 2012;2(3-5):186-201.https://doi.org/10.1159/000334260
[B125] Peça J, Feliciano C, Ting JT, 等人。Shank3 突变小鼠表现出类似自闭症的行为和纹状体功能障碍。自然。2011;472(7344):437-442。.https://doi.org/10.1038/nature09965
[B126] 申元和,李惠荣,纪贤惠,等. 恢复NMDA受体功能可改善SHANK2突变小鼠的孤独症样社交行为. Nature. 2012;486(7402):261-265.https://doi.org/10.1038/nature11208
[B127] Baudouin SJ, Gaudias J, Gerharz S, 等. 综合征和非综合征自闭症啮齿动物模型中共有的突触病理生理学. Science. 2012;338(6103):128-132.https://doi.org/10.1126/science.1224159
[B128] Huber KM, Gallagher SM, Warren ST, Bear MF. 脆性X智力迟钝小鼠模型中突触可塑性的改变。Proc Natl Acad Sci U S A。2002;99(11):7746-7750。.https://doi.org/10.1073/pnas.122205699
[B129] Ecker C, Bookheimer SY, Murphy DG. 神经影像学在自闭症谱系障碍中的应用:贯穿生命周期的脑结构与功能。柳叶刀神经病学。2015;14(11):1121-1134。.https://doi.org/10.1016/S1474-4422(15 )00050-2
[B130] Hazlett HC, Gu H, Munsell BC 等人。自闭症谱系障碍高风险婴儿早期大脑发育。自然。2017;542(7641):348-351。.https://doi.org/10.1038/nature21369
[B131] Miller EK, Cohen JD. 前额叶皮层功能整合理论。年度神经科学评论。2001;24:167-202。.https://doi.org/10.1146/annurev.neuro.24.1.167
[B132] Castelli F, Frith C, Happé F, Frith U. 自闭症、阿斯伯格综合征以及将心理状态归因于动态形状的大脑机制。《Brain》。2002;125(Pt 8):1839-1849。.https://doi.org/10.1093/brain/awf189
[B133] Just MA, Cherkassky VL, Keller TA, Kana RK, Minshew NJ. 功能和解剖上的大脑皮层连接不足与自闭症:一项执行功能任务的FMRI研究和胼胝体形态测量学证据。Cereb Cortex. 2007;17(4):951-961。.https://doi.org/10.1093/cercor/bhl006
[B134] Casanova MF,Buxhoeveden DP,Switala AE,Roy E。自闭症中的微柱病理学。神经学。2002;58(3):428-432。.https://doi.org/10.1212/wnl.58.3.428
[B135] Adolphs R. The social brain: neural basis of social knowledge. Annu Rev Psychol. 2009;60:693-716.https://doi.org/10.1146/annurev.psych.60.110707.163514
[B136] Nordahl CW, Scholz R, Yang X, 等. 谱系障碍儿童2至4岁时杏仁核生长速度加快:一项纵向研究. Arch Gen Psychiatry. 2012;69(1):53-61.https://doi.org/10.1001/archgenpsychiatry.2011.145
[B137] Kleinhans NM, Richards T, Sterling L 等人. 自闭症谱系障碍在面部加工过程中的功能连接异常。《Brain》。2008;131(Pt 4):1000-1012。.https://doi.org/10.1093/brain/awm334
[B138] Monk CS、Weng SJ、Wiggins JL 等人。自闭症谱系障碍中情绪面部处理的神经回路。J Psychiatry Neurosci。2010;35(2):105-114。.https://doi.org/10.1503/jpn.090085
[B139] Schmahmann JD. 小脑与认知。神经科学快报。2019;688:62-75。.https://doi.org/10.1016/j.neulet.2018.07.005
[B140] Fatemi SH, Aldinger KA, Ashwood P, 等人。共识论文:小脑在自闭症中的病理作用。小脑。2012;11(3):777-807。.https://doi.org/10.1007/s12311-012-0355-9
[B141] Courchesne E, Yeung-Courchesne R, Press GA, Hesselink JR, Jernigan TL. 孤独症患者小脑蚓叶VI和VII发育不全。《新英格兰医学杂志》,1988;318(21):1349-1354。.https://doi.org/10.1056/NEJM198805263182102
[B142] Wang SS, Kloth AD, Badura A. 小脑、敏感期和自闭症. Neuron. 2014;83(3):518-532.https://doi.org/10.1016/j.neuron.2014.07.016
[B143] Squire LR, Stark CE, Clark RE. 内侧颞叶。年神经科学评论。2004;27:279-306。.https://doi.org/10.1146/annurev.neuro.27.070203.144130
[B144] Schumann CM, Hamstra J, Goodlin-Jones BL, et al. 自闭症儿童杏仁核增大,青少年则不然;海马体在所有年龄段都增大。J Neurosci. 2004;24(28):6392-6401。.https://doi.org/10.1523/JNEUROSCI.1297-04.2004
[B145] Gaigg SB, Bowler DM。自闭症谱系障碍中情绪唤起词的自由回忆和遗忘。神经心理学。2008;46(9):2336-2343。.https://doi.org/10.1016/j.neuropsychologia.2008.03.008
[B146] Graybiel AM. 基底神经节. Curr Biol. 2000;10(14):R509-R511.https://doi.org/10.1016/s0960-9822(00 )00593-5
[B147] Langen M, Durston S, Staal WG, Palmen SJ, van Engeland H. 孤独症高功能未用药受试者尾状核增大。Biol Psychiatry. 2007;62(3):262-266。.https://doi.org/10.1016/j.biopsych.2006.09.040
[B148] Scott-Van Zeeland AA, Dapretto M, Ghahremani DG, Poldrack RA, Bookheimer SY. 40%的自闭症患者接受奖励处理。自闭症研究。2010;3(2):53-67。.https://doi.org/10.1002/aur.122
[B149] Gershenzon J, Dudareva N. 萜烯天然产物在自然界中的功能. Nat Chem Biol. 2007;3(7):408-414.https://doi.org/10.1038/nchembio.2007.5
[B150] Russo EB。驯服 THC:潜在的大麻协同作用和植物大麻素-萜烯的伴随效应。Br J Pharmacol。2011;163(7):1344-1364。.https://doi.org/10.1111/j.1476-5381.2011.01238.x
[B151] Aprotosoaie AC, Hăncianu M, Costache II, Miron A. 芳樟醇:一种具有宝贵生物特性的关键气味分子的综述。Flavour Fragr J. 2014;29(4):193-219。.https://doi.org/10.1002/ffj.3197
[B152] 原田浩, 柏谷宏, 神村由美, 桑木崇. 芳樟醇气味诱导的小鼠抗焦虑作用. Front Behav Neurosci. 2018;12:241.https://doi.org/10.3389/fnbeh.2018.00241
[B153] 埃利萨贝茨基 E,布鲁姆 LF,德索萨 DO。 在谷氨酸相关癫痫模型中 Linalool 的抗惊厥特性。植物医学。1999;6(2):107-113。.https://doi.org/10.1016/S0944-7113(99 )80044-0
[B154] Linck VM, da Silva AL, Figueiró M, 等人。吸入芳樟醇对小鼠焦虑、社交互动和攻击行为的影响。Phytomedicine。2010;17(8-9):679-683。.https://doi.org/10.1016/j.phymed.2009.10.002
[B155] Takahashi M, Satou T, Ohashi M, Hayashi S, Sadamoto K, Koike K. 薰衣草精油的种间化学成分比较及吸入镇静作用. Nat Prod Commun. 2011;6(11):1769-1774.https://journals.sagepub.com/doi/10.1177/1934578X1100601148
[B156] Peana AT, D’Aquila PS, Panin F, Serra G, Pippia P, Moretti MD. 芳樟醇和芳樟醇乙酸酯在精油中的抗炎活性。植物药学。2002;9(8):721-726。.https://doi.org/10.1078/094471102321621322
[B157] Sabogal-Guáqueta AM, Posada-Duque R, Cortes NC, Arias-Londoño JD, Cardona-Gómez GP. 海马体和外周磷脂谱的变化与长期全球性脑缺血模型中神经退行性变标志物相关:芳樟醇的减轻作用。神经药理学。2018;135:555-571。.https://doi.org/10.1016/j.neuropharm.2018.04.015
[B158] Jäger W, Buchbauer G, Jirovetz L, Fritzer M. 薰衣草油作为按摩油的经皮吸收. J Soc Cosmet Chem. 1992;43:49-54.https://www.researchgate.net/publication/232711524_Percutaneous_absorption_of_lavender_oil_from_a_massage_oil
[B159] Buchbauer G. 挥发油的生物活性. 载: Baser KHC, Buchbauer G, 主编. 挥发油手册:科学、技术与应用. 第2版. CRC Press; 2015:235-280.https://www.routledge.com/Handbook-of-Essential-Oils-Science-Technology-and-Applications/Baser-Buchbauer/p/book/9781032046419
[B160] Gertsch J, Leonti M, Raduner S, et al. β-石竹烯是一种膳食大麻素。Proc Natl Acad Sci U S A. 2008;105(26):9099-9104.https://doi.org/10.1073/pnas.0803601105
Bahi A, Al Mansouri S, Al Memari E, Al Ameri M, Nurulain SM, Ojha S. β-Caryophyllene, a CB2 receptor agonist produces multiple behavioral changes relevant to anxiety and depression in mice. Physiol Behav. 2014;135:119-124.https://doi.org/10.1016/j.physbeh.2014.06.003
[B162] Atwood BK, Mackie K. CB2:一个身份认同危机的大麻素受体。英属药理学杂志。2010;160(3):467-479。.https://doi.org/10.1111/j.1476-5381.2010.00729.x
[B163] Klauke AL, Racz I, Pradier B, 等人。 the cannabinoid CB₂ receptor-selective phytocannabinoid beta-caryophyllene exerts analgesic effects in mouse models of inflammatory and neuropathic pain. 欧洲神经精神药理学杂志。 2014;24(4):608-620。.https://doi.org/10.1016/j.euroneuro.2013.10.008
[B164] 程Y, 董Z, 刘S. β-石杉碱通过CB2受体激活和PPARγ通路改善APP/PS1小鼠的阿尔茨海默病样表型。《药理学》. 2014;94(1-2):1-12。.https://doi.org/10.1159/000362689
[B165] Askari VR, Shafiee-Nick R. β-石竹烯对LPS诱导的少突胶质细胞毒性有前景的神经保护作用:一项机制研究。Biochem Pharmacol. 2019;159:154-171。.https://doi.org/10.1016/j.bcp.2018.12.001
[B166] Fontes LBA, Dias D, Aarestrup BJ, et al. β-石竹烯改善C57BL/6小鼠实验性自身免疫性脑脊髓炎的发生。Biomed Pharmacother。2017;91:257-264。.https://doi.org/10.1016/j.biopha.2017.04.092
[B167] Alberti TB, Barbosa WL, Vieira JL, Raposo NR, Dutra RC. (-)-β-Caryophyllene,一种选择性作用于CB2受体的植物大麻素,在小鼠多发性硬化症模型中能抑制运动瘫痪和神经炎症。Int J Mol Sci. 2017;18(4):691.https://doi.org/10.3390/ijms18040691
[B168] Galdino PM, Nascimento MV, Florentino IF, 等. 源自 Spiranthera odoratissima A. St. Hil. 叶子的精油及其主要成分 β-石竹烯对雄性小鼠的抗焦虑样作用. Prog Neuropsychopharmacol Biol Psychiatry. 2012;38(2):276-284.https://doi.org/10.1016/j.pnpbp.2012.04.012
[B169] Horváth B, Mukhopadhyay P, Kechrid M 等人。β-石竹烯以大麻素 2 受体依赖性方式改善顺铂诱导的肾毒性。Free Radic Biol Med. 2012;52(8):1325-1333。.https://doi.org/10.1016/j.freeradbiomed.2012.01.014
[B170] Fidyt K, Fiedorowicz A, Strządała L, Szumny A. β-香叶烯和 β-香叶烯氧化物-具有抗癌和镇痛特性的天然化合物。Cancer Med. 2016;5(10):3007-3017。.https://doi.org/10.1002/cam4.816
[B171] Sharma C, Al Kaabi JM, Nurulain SM, Goyal SN, Kamal MA, Ojha S. β-香叶烯的药理作用及其治疗潜力:一种具有药用前景的膳食植物大麻素。Curr Pharm Des。2016;22(21):3237-3264。.https://doi.org/10.2174/1381612822666160311115226
[B172] Ammon HP. 乳香酸及其在慢性炎症性疾病中的作用。Adv Exp Med Biol。2016;928:291-327。.https://doi.org/10.1007/978-3-319-41334-1_13
[B173] Catanzaro D, Rancan S, Orso G, et al. 乳香(Boswellia serrata)保护肠道上皮屏障免受氧化和炎症损伤。PLoS One。2015;10(5):e0125375。.https://doi.org/10.1371/journal.pone.0125375
[B174] Khoshnazar M, Bigdeli MR, Parvardeh S, Pouriran R. α-蒎烯对大鼠局灶性缺血性中风引起的神经行为缺陷、氧化损伤和炎症反应的减弱作用。J Pharm Pharmacol. 2019;71(11):1725-1733。.https://doi.org/10.1111/jphp.13164
[B175] Ryter SW, Alam J, Choi AM. 产铁血红素加氧酶-1/一氧化碳:从基础科学到治疗应用。Physiol Rev. 2006;86(2):583-650。.https://doi.org/10.1152/physrev.00011.2005
[B176] Umar S, Umar K, Sarwar AH, 等. Boswellia serrata 提取物可减轻胶原诱导关节炎中的炎症介质和氧化应激。Phytomedicine。2014;21(6):847-856。.https://doi.org/10.1016/j.phymed.2014.02.001
[B177] Abdel-Tawab M, Werz O, Schubert-Zsilavecz M. 印度乳香:体外、临床前、药代动力学和临床数据的整体评估。Clin Pharmacokinet. 2011;50(6):349-369。.https://doi.org/10.2165/11586800-000000000-00000
[B178] Hosseini M, Hadjzadeh MA, Derakhshan M, et al. 乳香对成年大鼠甲状腺功能减退引起的记忆缺陷在莫里斯水迷宫中的有益作用. Arch Pharm Res. 2010;33(3):463-468.https://doi.org/10.1007/s12272-010-0316-6
[B179] Safayhi H, Mack T, Sabieraj J, Anazodo MI, Subramanian LR, Ammon HP. 没药酸:新型、特异性、非氧化还原的5-脂氧合酶抑制剂。J Pharmacol Exp Ther。1992;261(3):1143-1146。.https://www.sciencedirect.com/science/article/abs/pii/S0022356525111300
[B180] 维尔茨·奥. 5-脂氧合酶:细胞生物学和分子药理学。Curr Drug Targets Inflamm Allergy。2002;1(1):23-44。.https://doi.org/10.2174/1568010023344959
[B181] Al-Harrasi A, Ali L, Hussain J,等。阿曼乳香粗提物和馏分镇痛作用,来自传统药用植物乳香树 Boswellia sacra,在动物模型上。Asian Pac J Trop Med。2014;7S1:S485-S490。.https://doi.org/10.1016/S1995-7645(14 )60279-0
[B182] Cimanga K, Kambu K, Tona L, 等. 刚果民主共和国部分芳香药用植物精油的化学成分与抗菌活性相关性研究. J Ethnopharmacol. 2002;79(2):213-220.https://doi.org/10.1016/s0378-8741(01 )00384-1
[B183] 佐藤T,春屋K,前田K,小池K。小鼠日常吸入α-蒎烯:对行为和器官积累的影响。植疗研究。2014;28(9):1284-1287。.https://doi.org/10.1002/ptr.5105
[B184] 佐藤(Satou)T, 村上(Murakami)S, 松浦(Matsuura)M, 林(Hayashi)S, 小池(Koike)K. 吸入式艳山姜(Alpinia zerumbet)精油在小鼠中的抗焦虑作用和组织分布. Nat Prod Commun. 2010;5(1):143-146.https://journals.sagepub.com/doi/10.1177/1934578X1000500133
[B185] Juergens UR, Dethlefsen U, Steinkamp G, Gillissen A, Repges R, Vetter H. 1.8-桉叶油素(桉叶油醇)在支气管哮喘中的抗炎活性:一项双盲安慰剂对照试验。Respir Med. 2003;97(3):250-256。.https://doi.org/10.1053/rmed.2003.1432
[B186] Santos FA, Rao VS. 1,8-桉叶油素的抗炎和镇痛作用——一种存在于多种植物精油中的萜类氧化物。植物疗法研究。2000;14(4):240-244。.https://doi.org/10.1002/1099-1573(200006 3.0.co;2-x
[B187] Tisserand R, Young R. 精油安全:保健专业人士指南。第 2 版。丘吉尔·利文斯顿;2014 年。.https://www.elsevier.com/books/essential-oil-safety/tisserand/978-0-7020-4606-4
[B188] Worth H, Schacher C, Dethlefsen U. 桉叶油 (桉叶素) 联用治疗可减少 COPD exacerbations:一项安慰剂对照双盲试验。Respir Res. 2009;10:69。.https://doi.org/10.1186/1465-9921-10-69
[B189] Salehi B, Upadhyay S, Erdogan Orhan I, et al. α-和β-蒎烯的治疗潜力:大自然的奇迹馈赠。Biomolecules。2019;9(11):738。.https://doi.org/10.3390/biom9110738
[B190] Guzmán-Gutiérrez SL, Gómez-Cansino R, García-Zebadúa JC, Jiménez-Pérez NC, Reyes-Chilpa R. Litsea glaucescens 精油的抗抑郁活性:以 β-蒎烯和芳樟醇为活性成分的鉴定。《民族药理学杂志》,2012;143(2):673-679。.https://doi.org/10.1016/j.jep.2012.07.026
[B191] 杨 H, 禹 J, 裴 AN, 等. pine tree oils 的主要成分 α-蒎烯通过 GABAA-苯二氮䓬受体增强小鼠非快速眼动睡眠. Mol Pharmacol. 2016;90(5):530-539.https://doi.org/10.1124/mol.116.105080
[B192] Kim DS, Lee HJ, Jeon YD, 等人。 α-蒎烯通过抑制小鼠腹膜巨噬细胞的MAPK和NF-κB通路表现出抗炎活性。 Am J Chin Med。 2015;43(4):731-742。.https://doi.org/10.1142/S0192415X15500457
[B193] Porres-Martínez M, González-Burgos E, Carretero ME, Gómez-Serranillos MP。 狭叶鼠尾草精油抗氧化应激诱导的神经损伤的保护作用。 Food Chem Toxicol。 2015;80:154-162。.https://doi.org/10.1016/j.fct.2015.03.002
[B194] 宫泽美佐子、渡边浩、龟冈弘。对具有对薄荷烷骨架的单萜抑制乙酰胆碱酯酶活性的研究。J Agric Food Chem。1997;45(3):677-679。.https://doi.org/10.1021/jf960398b
[B195] Deutsch SI, Burket JA, Katz E. 神经元迁移的微小紊乱是否会加剧精神分裂症和其他神经发育障碍?潜在的遗传机制及可能的治疗启示。Eur Neuropsychopharmacol. 2010;20(5):281-287。.https://doi.org/10.1016/j.euroneuro.2010.02.005
[B196] Tan LT, Lee LH, Yin WF, et al. 依兰(Cananga odorata)的传统用途、植物化学和生物活性。Evid Based Complement Alternat Med. 2015;2015:896314.https://doi.org/10.1155/2015/896314
[B197] Jung YH, Ha TM, Ahn J, et al. 依兰依兰精油吸入改善丙戊酸诱导的自闭症谱系障碍大鼠的社交互动和焦虑样行为。Nutrients。2021;13(11):3804。.https://doi.org/10.3390/nu13113804
[B198] Sinha Y, Silove N, Wheeler D, Williams K. 听觉整合训练和其他声音疗法在自闭症谱系障碍中的应用:系统性回顾。《英国儿科学杂志》。2006;91(12):1018-1022。.https://doi.org/10.1136/adc.2006.094649
[B199] Cullen-Powell LA, Barlow JH, Cushway D. 探索为患有自闭症的儿童及其父母提供的按摩干预措施:对亲子关系和依恋的影响。《儿童健康杂志》。2005;9(4):245-255。.https://doi.org/10.1177/1367493505056479
[B200] Buckle J. 临床芳香疗法:精油在医疗保健中的应用. 第3版. Churchill Livingstone; 2014.https://www.elsevier.com/books/clinical-aromatherapy/buckle/978-0-7020-5440-3
[B201] Saiyudthong S, Marsden CA. 佛手柑精油对大鼠焦虑相关行为和皮质酮水平的急性效应。植物药研究。2011;25(6):858-862。.https://doi.org/10.1002/ptr.3325
[B202] Navarra M, Mannucci C, Delbò M, Calapai G. 香柠檬精油:从基础研究到临床应用。Front Pharmacol. 2015;6:36。.https://doi.org/10.3389/fphar.2015.00036
[B203] Efferth T, Oesch F. 乳香的抗炎和抗癌活性:靶点、治疗和毒性。Semin Cancer Biol。2022;80:39-57。.https://doi.org/10.1016/j.semcancer.2020.01.015
[B204] Siddiqui MZ. 乳香(Boswellia serrata),一种潜在的抗炎剂:概述。印度药学杂志。2011;73(3):255-261。.https://doi.org/10.4103/0250-474X.93507
[B205] Hamidpour R, Hamidpour S, Hamidpour M, Shahlari M. 乳香(Rǔ xiāng;乳香属):从传统应用选择到预防和治疗严重疾病的新型植物疗法。J Tradit Complement Med. 2013;3(4):221-226。.https://doi.org/10.4103/2225-4110.119723
[B206] Moussaieff A, Mechoulam R. 搏斯韦利亚树脂:从宗教仪式到医疗用途;一项体外、体内和临床试验的综述。J Pharm Pharmacol。2009;61(10):1281-1293。.https://doi.org/10.1211/jpp/61.10.0003
[B207] Bakkali F, Averbeck S, Averbeck D, Idaomar M. 具有生物活性的精油——一项综述。食品化学毒理学。2008;46(2):446-475。.https://doi.org/10.1016/j.fct.2007.09.106
[B208] Setzer WN. 精油与抗焦虑芳香疗法。Nat Prod Commun。2009;4(9):1305-1316。.https://pubmed.ncbi.nlm.nih.gov/19831048/
[B209] Adams RP. 气相色谱/质谱法鉴定精油成分。第4版。Allured Publishing;2007。.https://www.allured.com/identification-of-essential-oil-components-by-gas-chromatographymass-spectrometry-4th-edition
[B210] Dayawansa S, Umeno K, Takakura H, 等人. 沉香醇天然香味吸入对人体自主神经反应的影响. Auton Neurosci. 2003;108(1-2):79-86.https://doi.org/10.1016/j.autneu.2003.08.002
[B211] Cheaha D, Reakkamnuan C, Nukitram J, et al. 影响睡眠-觉醒和脑电图的岩兰草精油吸入的改良。J Intercult Ethnopharmacol. 2016;5(1):72-78。.https://doi.org/10.5455/jice.20160208050736
[B212] Friedmann T. 摆脱ADHD:儿童ADHD的无药物治疗方法. Lotus Press; 2000.https://www.researchgate.net/publication/265127532_Attention_Deficit_and_Hyperactivity_Disorder_ADHD
[B213] Weyerstahl P, Marschall H, Weirauch M, Thefeld K, Surburg H. 商业香根草油的成分。Flavour Fragr J。2000;15(3):153-173。.https://doi.org/10.1002/1099-1026(200005/06 )15:33.0.CO;2-V
[B214] 普里普迪维奇 P,春蓬西里 W,苏蒂阿波恩 P,黄鹏猜 S。泰国罗勒的化学成分和抗氧化活性,采用保留指数和二维气相色谱联用技术。J Serb Chem Soc。2010;75(11):1503-1513。.https://doi.org/10.2298/JSC100203125P
[B215] Hongratanaworakit T, Buchbauer G. 依兰油经皮吸收后对人体的放松作用。Phytother Res. 2006;20(9):758-763。.https://doi.org/10.1002/ptr.1950
[B216] 莫斯 M,休伊特 S,莫斯 L,韦斯内斯 K。薄荷和依兰香气对认知表现和情绪的调节。Int J Neurosci。2008;118(1):59-77。.https://doi.org/10.1080/00207450601042094
[B217] 小宮満、竹内忠、原田英明。柠檬精油蒸汽通过调节小鼠5-HT和DA活性产生抗应激作用。行为脑研究。2006;172(2):240-249。.https://doi.org/10.1016/j.bbr.2006.05.006
[B218] Venturi GB. 关于流体横向运动原理的实验研究。英国皇家学会哲学汇刊。1797;87:243-260。.https://royalsocietypublishing.org/doi/10.1098/rstl.1799.0005
[B219] Buckle J. 芳香疗法作为慢性疼痛辅助治疗的应用。Altern Ther Health Med。1999;5(5):42-51。.https://pubmed.ncbi.nlm.nih.gov/10450129/
[B220] Hongratanaworakit T. 按摩精油混合物对人体的芳香疗法作用。《天然产物杂志》。2011;6(8):1199-1204。.https://www.researchgate.net/publication/51647617_Aroma-therapeutic_Effects_of_Massage_Blended_Essential_Oils_on_Humans
[B221] 怀特 FM. 流体机械. 第8版. 麦格劳-希尔教育出版社; 2015.https://www.mheducation.com/highered/product/fluid-mechanics-white/M9781260258318.html
[B222] Haze S, Sakai K, Gozu Y. 香气吸入对正常成人交感神经活动的影.https://doi.org/10.1254/jjp.90.247
[B223] Herz RS. 香薰疗法的事实与虚构:对嗅觉对情绪、生理和行为影响的科学分析。国际神经科学杂志。2009;119(2):263-290。.https://doi.org/10.1080/00207450802333953
[B224] Turek C, Stintzing FC. 精油的稳定性:一篇综述. Comprehensive Reviews in Food Science and Food Safety. 2013;12(1):40-53.https://doi.org/10.1111/1541-4337.12006
[B225] Buckle J. 临床芳香疗法在护理中的应用. 第2版. 整体护理实践; 2001.https://journals.lww.com/holistic-nursing-practice/citation/2001/04000/clinical_aromatherapy_in_nursing.15.aspx
[B226] Labiris NR, Dolovich MB. 肺部药物输送。第一部分:影响气溶胶药物治疗效果的生理因素。英国临床药理学杂志。2003;56(6):588-599。.https://doi.org/10.1046/j.1365-2125.2003.01892.x
[B227] Price S, Price L. 芳香疗法在保健专业人员中的应用. 第4版. 丘吉尔·利文斯顿; 2011.https://www.elsevier.com/books/aromatherapy-for-health-professionals/price/978-0-7020-3151-0
[B228] Battaglia S. 芳香疗法全指南. 第3版. Black Pepper Creative; 2018.https://www.blackpeppercreative.com.au/the-complete-guide-to-aromatherapy
[B229] Posadzki P, Alotaibi A, Ernst E. 芳香疗法的副作用:病例报告和病例系列系统评价。《国际风险与安全医学杂志》2012;24(3):147-161。.https://doi.org/10.3233/JRS-2012-0568
[B230] Dunn C, Sleep J, Collett D. Sensing an improvement: an experimental study to evaluate the use of aromatherapy, massage and periods of rest in an intensive care unit. J Adv Nurs. 1995;21(1):34-40.https://doi.org/10.1046/j.1365-2648.1995.21010034.x
[B231] Bowles EJ. aromatherapeutic oils的化学. 第3版. Allen & Unwin; 2003.https://www.allenandunwin.com/browse/books/general-books/health-wellbeing/The-Chemistry-of-Aromatherapeutic-Oils-E-Joy-Bowles-9781741140217
[B232] Schnaubelt K. 高级芳香疗法:精油疗法的科学。Healing Arts Press;1998。.https://www.healingartspressbooks.com/advanced-aromatherapy
[B233] Schaaf RC, Lane AE. Toward a best-practice protocol for assessment of sensory features in ASD. J Autism Dev Disord. 2015;45(5):1380-1395.https://doi.org/10.1007/s10803-014-2299-z
[B234] Wolkoff P, Wilkins CK, Clausen PA, Nielsen GD. 办公室环境中的有机化合物——感官刺激、气味、测量以及反应化学的作用。室内空气。2006;16(1):7-19。.https://doi.org/10.1111/j.1600-0668.2005.00393.x
[B235] Dalton P. 嗅觉适应与交叉适应。载:Doty RL,编。嗅觉与味觉手册。第3版。Wiley-Blackwell;2015:443-459。.https://www.wiley.com/en-us/Handbook+of+Olfaction+and+Gustation%2C+3rd+Edition-p-9780470455951
[B236] Cain WS. 用鼻子去认识:气味识别的关键. 科学. 1979;203(4379):467-470.https://doi.org/10.1126/science.760202
[B237] Kern JK, Trivedi MH, Garver CR, et al. autismsense processing disorder in children. Autism. 2006;10(5):480-494.https://doi.org/10.1177/1362361306066564
[B238] Lehrner J, Marwinski G, Lehr S, Johren P, Deecke L. 橙子和薰衣草的室内气味能缓解焦虑并改善牙科诊所里的情绪。《生理行为学》杂志。2005;86(1-2):92-95。.https://doi.org/10.1016/j.physbeh.2005.06.031
[B239] Cavanagh HM, Wilkinson JM. 薰衣草精油的生物活性。植物疗法研究。2002;16(4):301-308。.https://doi.org/10.1002/ptr.1103
[B240] Diego MA, Jones NA, Field T, 等人。芳香疗法对情绪、警觉性脑电图模式和数学运算有积极影响。Int J Neurosci。1998;96(3-4):217-224。.https://doi.org/10.3109/00207459808986469
[B241] 田野T,迭戈M,赫尔南德斯-赖夫M,西斯内罗斯W,费霍L,维拉Y,吉尔K,格林纳D,克莱尔何Q。薰衣草香味洁面凝胶对放松的影响。《国际神经科学杂志》。2005;115(2):207-222。.https://doi.org/10.1080/00207450590519175
[B242] Goel N, Kim H, Lao RP. 嗅觉刺激可改变年轻男女的夜间睡眠。《生物钟国际》. 2005;22(5):889-904。.https://doi.org/10.1080/07420520500263276
[B243] Ashburner J, Ziviani J, Rodger S. 自闭症谱系障碍儿童的感官处理与课堂情绪、行为及教育结局. Am J Occup Ther. 2008;62(5):564-573.https://doi.org/10.5014/ajot.62.5.564
[B244] Tomchek SD, Dunn W. 儿童和自闭症儿童的感官处理:一项使用简易感官特征的比较研究。Am J Occup Ther. 2007;61(2):190-200。.https://doi.org/10.5014/ajot.61.2.190
[B245] Ben-Sasson A, Hen L, Fluss R, Cermak SA, Engel-Yeger B, Gal E. 自闭症谱系障碍个体感觉调节症状的荟萃分析。《自闭症与发育障碍杂志》。2009;39(1):1-11。.https://doi.org/10.1007/s10803-008-0593-3
Leekam SR, Nieto C, Libby SJ, Wing L, Gould J. 描述自闭症儿童和成人的感觉异常。J Autism Dev Disord. 2007;37(5):894-910.https://doi.org/10.1007/s10803-006-0218-7
[B247] Baranek GT, David FJ, Poe MD, Stone WL, Watson LR. 感官体验问卷:区分自闭症、发育迟缓和正常发育的幼儿的感官特征。《儿童心理学与精神病学杂志》. 2006;47(6):591-601。.https://doi.org/10.1111/j.1469-7610.2005.01546.x
[B248] Herz RS. 嗅觉诱发记忆在心理和生理健康中的作用。Brain Sci. 2016;6(3):22。.https://doi.org/10.3390/brainsci6030022
[B249] Mesibov GB, Shea V, Schopler E. 孤独症谱系障碍的TEACCH方法。Springer出版社;2004年。.https://link.springer.com/book/10.1007/978-0-387-23855-2
[B250] 大野IP、Honey EJ、McConachie H。针对自闭症谱系障碍(ASD)幼儿的家长媒介早期干预。Cochrane数据库系统评价。2013;(4):CD009774。.https://doi.org/10.1002/14651858.CD009774.pub2
[B251] Marriott PJ, Shellie R, Cornwell C. 气相色谱技术在精油分析中的应用. J Chromatogr A. 2001;936(1-2):1-22.https://doi.org/10.1016/s0021-9673(01 )01314-0
[B252] Sparkman OD, Penton Z, Kitson FG. 气相色谱与质谱:实用指南。第2版。Academic Press;2011。.https://www.sciencedirect.com/book/9780123736284/gas-chromatography-and-mass-spectrometry
[B253] ISO 4720:2009. 精油 – 命名. 国际标准化组织; 2009.https://www.iso.org/standard/41076.html
[B254] ISO 11024-1:1998. 精油 – 色谱图一般指导 – 第1部分:色谱图在标准中的准备. 国际标准化组织;1998年.https://www.iso.org/standard/19053.html
[B255] Dhifi W, Bellili S, Jazi S, Bahloul N, Mnif W. 精油的化学特征及其部分生物活性的研究:一篇批判性综述. Medicines (Basel). 2016;3(4):25.https://doi.org/10.3390/medicines3040025
[B256] 莉斯-巴尔钦·M. 芳香疗法科学:医疗保健专业人员指南. 药学出版社;2006。.https://www.pharmpress.com/product/9780853696230/aromatherapy-science
[B257] Schnaubelt K. 芳香疗法医学:精油疗法。Frog Books;1999。.https://www.innertraditions.com/books/medical-aromatherapy
[B258] Tisserand R. 芳香疗法艺术.healing arts press; 1977.https://www.innertraditions.com/books/the-art-of-aromatherapy
[B259] Pauli A. 精油成分的抗菌性能。《国际芳香疗法杂志》。2001;11(3):126-133。.https://doi.org/10.1016/S0962-4562(01 )80048-5
欧洲药典委员会。欧洲药典。第10版。欧洲委员会;2019。.https://www.edqm.eu/en/european-pharmacopoeia-ph-eur-11th-edition
[B261] Wabner D, Beier C. 芳香疗法:基础、作用原理、实践。第2版。Urban & Fischer;2012年。.https://www.elsevier.com/books/aromatherapie/wabner/978-3-437-57214-4
[B262] Lis-Balchin M. 天竺葵与香叶天竺葵:天竺葵属和香叶天竺葵属。CRC出版社;2002年。.https://www.routledge.com/Geranium-and-Pelargonium/Lis-Balchin/p/book/9780415284653
[B263] Turek C, Stintzing FC. 精油的稳定性:综述。Compr Rev Food Sci Food Saf。2013;12(1):40-53。.https://doi.org/10.1111/1541-4337.12006
[B264] Skold M, Karlberg AT, Matura M, Börje A. 芳香化学品β-石竹烯–空气氧化和皮肤致敏。食品化学毒理学。2006;44(4):538-545。.https://doi.org/10.1016/j.fct.2005.08.028
[B265] Tavassoli T, Miller LJ, Schoen SA, Nielsen DM, Baron-Cohen S. 成人自闭症谱系障碍人群的感官过度反应. Autism. 2014;18(4):428-432.https://doi.org/10.1177/1362361313477246
[B266] Lane AE, Young RL, Baker AE, Angley MT. 自闭症中的感觉加工亚型:与适应性行为的关系。《自闭症与发育障碍杂志》。2010;40(1):112-122。.https://doi.org/10.1007/s10803-009-0840-2
[B267] Mazurek MO, Vasa RA, Kalb LG, 等人。孤独症谱系障碍儿童的焦虑、感觉过敏和胃肠道问题。J Abnorm Child Psychol。2013;41(1):165-176。.https://doi.org/10.1007/s10802-012-9668-x
[B268] Buckle J. 临床芳香疗法:精油实践. 第2版. Churchill Livingstone; 2003.https://www.elsevier.com/books/clinical-aromatherapy/buckle/978-0-7020-2536-6
[B269] Tuchman R, Cuccaro M, Alessandri M. 自闭症与癫痫:历史视角. Brain Dev. 2010;32(9):709-718.https://doi.org/10.1016/j.braindev.2010.04.008
[B270] Burkhard PR, Burkhardt K, Haenggeli CA, Landis T. 植物引起的癫痫发作:一个老问题再次出现。J Neurol. 1999;246(8):667-670。.https://doi.org/10.1007/s004150050434
[B271] Millet Y, Jouglard J, Steinmetz MD, Tognetti P, Joanny P, Arditti J. 一些必需植物精油的毒性。临床和实验研究。Clin Toxicol. 1981;18(12):1485-1498.https://doi.org/10.3109/15563658108990111
[B272] Pengelly A, Bennett K, Spinks J, Bensoussan A. 评价使用精油治疗癫痫的证据基础:一项系统评价。Epilepsy Behav. 2012;24(1):9-15。.https://doi.org/10.1016/j.yebeh.2012.02.019
[B273] Höld KM, Sirisoma NS, Ikeda T, Narahashi T, Casida JE。α-侧柏酮(苦艾酒的活性成分):γ-氨基丁酸A型受体调节和代谢解毒。Proc Natl Acad Sci U S A。2000;97(8):3826-3831。.https://doi.org/10.1073/pnas.070042397
[B274] Millet Y, Tognetti P, Lavaire-Perlovisi M, Steinmetz MD, Arditti J, Jouglard J。关于牛至和鼠尾草精油商业样品兴奋特性的研究。Plantes Med Phytother。1979;13(3):179-184。.https://www.sciencedirect.com/science/article/abs/pii/S0370447579800477
[B275] Marcus C, Lichtenstein EP. 天然的食物植物成分与杀虫剂和戊巴比妥在大鼠和小鼠中的相互作用。J Agric Food Chem. 1979;27(5):1217-1223。.https://doi.org/10.1021/jf60226a004
[B276] Koutroumanidou E, Kimbaris A, Kortsaris A, 等. 经香精油给药后,戊四氮诱导的小鼠癫痫发作潜伏期延长,发作严重程度降低. Epilepsy Res Treat. 2013;2013:532657.https://doi.org/10.1155/2013/532657
[B277] Avallone R, Zanoli P, Puia G, Kleinschnitz M, Schreier P, Baraldi M. 药理学研究:从洋甘菊中分离出的黄酮类化合物芹菜素. 生物化学与药理学. 2000;59(11):1387-1394.https://doi.org/10.1016/s0006-2952(00 )00264-1
[B278] 渡边E,库赫塔K,木村M,劳瓦尔德HW,龟井T,今西J。佛手柑(芸香科柑橘属(Risso)Wright & Arn.)精油香薰对41名健康女性情绪状态、副交感神经系统活性及唾液皮质醇水平的影响。《德语传统医学研究》2015;22(1):43-49。.https://doi.org/10.1159/000380989
[B279] Ilhan S, Savaşer S, Kölgelier S. 芳香疗法对癫痫患者的影响. Epilepsy Behav. 2004;5(5):753-757.https://doi.org/10.1016/j.yebeh.2004.06.004
[B280] Meamarbashi A, Rajabi A. 薄荷对运动表现的影响. J Int Soc Sports Nutr. 2013;10(1):15.https://doi.org/10.1186/1550-2783-10-15
[B281] Eccles R. 薄荷醇及相关清凉化合物。J Pharm Pharmacol. 1994;46(8):618-630。.https://doi.org/10.1111/j.2042-7158.1994.tb03871.x
[B282] Silva J, Abebe W, Sousa SM, Duarte VG, Machado MI, Matos FJ. 桉树精油的镇痛和抗炎作用。民族药理学杂志。2003;89(2-3):277-283。.https://doi.org/10.1016/j.jep.2003.09.007
[B283] Fachini-Queiroz FC, Kummer R, Estevão-Silva CF, 等人。百里香酚和香芹酚(牛至精油成分)对炎症反应的影响。循证补充替代医学杂志。2012;2012:657026。.https://doi.org/10.1155/2012/657026
[B284] Carson CF, Hammer KA, Riley TV. 茶树油(Melaleuca alternifolia):抗菌及其他药用特性的回顾. Clin Microbiol Rev. 2006;19(1):50-62.https://doi.org/10.1128/CMR.19.1.50-62.2006
[B285] Eccles R, Jawad MS, Morris S. 口服施用(-)-薄荷醇对普通感冒引起的鼻塞患者气流鼻阻及鼻气流感觉的影响。J Pharm Pharmacol。1990;42(9):652-654。.https://doi.org/10.1111/j.2042-7158.1990.tb06625.x
[B286] Altunbas MK, Celık F, Kahraman C. 薰衣草精油在新生儿中安全使用的潜力。J Neonatal Nurs. 2020;26(6):303-307。.https://doi.org/10.1016/j.jnn.2020.05.005
[B287] Srivastava JK, Shankar E, Gupta S. 洋甘菊:一种过去时的草药,拥有光明的未来。Mol Med Rep。2010;3(6):895-901。.https://doi.org/10.3892/mmr.2010.377
[B288] Kiecolt-Glaser JK, Graham JE, Malarkey WB, Porter K, Lemeshow S, Glaser R. 嗅觉对情绪及自主神经、内分泌和免疫功能的影响. Psychoneuroendocrinology. 2008;33(3):328-339.https://doi.org/10.1016/j.psyneuen.2007.11.015
[B289] Halm MA. 精油在危重症患者症状管理中的应用。《美国危重监护杂志》。2008;17(2):160-163。.https://www.researchgate.net/publication/5541002_Essential_Oils_for_Management_of_Symptoms_in_Critically_Ill_Patients
[B290] Fung LK, Mahajan R, Nozzolillo A, 等. autism谱障碍的严重易激惹和问题行为的药物治疗:系统评价和荟萃分析. Pediatrics. 2016;137 Suppl 2:S124-S135.https://doi.org/10.1542/peds.2015-2851K
[B291] Kirschbaum KM, Müller MJ, Malevani J, et al. 血清阿立哌唑和脱氢阿立哌唑水平,临床反应和副作用。《世界生物精神病学杂志》。2008;9(3):212-218。.https://doi.org/10.1080/15622970701361255
[B292] Sprouse J, Clarke T, Reynolds L, Heym J, Rollema H. 舍曲林及其代谢产物去甲舍曲林在体内阻断中枢5-HT再摄取作用的比较。神经精神药物学。1996;14(4):225-231。.https://doi.org/10.1016/0893-133X(95 )00112-Q
[B293] Bailey DG, Dresser G, Arnold JM. 葡萄柚-药物相互作用:禁果还是可避免的后果?CMAJ。2013;185 (4):309-316。.https://doi.org/10.1503/cmaj.120951
[B294] Obach RS, Walsky RL, Venkatakrishnan K, Gaman EA, Houston JB, Tremaine LM. 体外细胞色素P450抑制数据在药物-药物相互作用预测中的应用。J Pharmacol Exp Ther。2006;316(1):336-348。.https://doi.org/10.1124/jpet.105.093229
[B295] 郭 L Q,山添 Y。葡萄柚汁和草药中香豆素对细胞色素 P450 的抑制作用。《中国药理学报》。2004;25(2):129-136。.https://www.researchgate.net/publication/8881220_Inhibition_of_cytochrome_P450_by_furanocoumarins_in_grapefruit_juice_and_herbal_medicines
[B296] Paine MF, Criss AB, Watkins PB. 两种主要的葡萄柚汁成分在肠道CYP3A4抑制作用开始时间上存在差异。J Pharmacol Exp Ther。2004;312(3):1151-1160。.https://doi.org/10.1124/jpet.104.076570
[B297] Hanley MJ, Cancalon P, Widmer WW, Greenblatt DJ. 葡萄柚汁对药物处置的影响。Expert Opin Drug Metab Toxicol. 2011;7(3):267-286.https://doi.org/10.1517/17425255.2011.553189
[B298] Dresser GK, Bailey DG. 饮用果汁对药物代谢的影响:药物相互作用的新模型。Eur J Clin Invest. 2003;33 Suppl 2:10-16。.https://doi.org/10.1046/j.1365-2362.33.s2.2.x
[B299] Izzo AA, Ernst E. 草药与处方药的相互作用:一项更新的系统评价。《药物》杂志。2009;69(13):1777-1798。.https://doi.org/10.2165/11317010-000000000-00000
[B300] Henderson L, Yue QY, Bergquist C, Gerden B, Arlett P. 圣约翰草(贯叶连翘):药物相互作用和临床结局。英国临床药理学杂志。2002;54(4):349-356。.https://doi.org/10.1046/j.1365-2125.2002.01683.x
[B301] Unger M, Frank A. 同时测定草药提取物对六种主要细胞色素P450酶活性的抑制效力,采用液相色谱/质谱联用和自动在线提取。Rapid Commun Mass Spectrom. 2004;18(19):2273-2281。.https://doi.org/10.1002/rcm.1621
[B302] Gurley BJ, Gardner SF, Hubbard MA, 等人。金印草、卡瓦卡瓦、黑升麻和缬草对人体细胞色素 P450 1A2、2D6、2E1 和 3A4/5 表型的影响。Clin Pharmacol Ther。2005;77(5):415-426。.https://doi.org/10.1016/j.clpt.2005.01.009
[B303] Spinella M. 精神活性草药药理协同作用的重要性。替代医学评论。2002;7(2):130-137。.https://pubmed.ncbi.nlm.nih.gov/11968843/
[B304] Abourashed EA, Koetter U, Brattström A. 缬草、啤酒花及其固定组合提取物(Ze91019)与选定中枢神经系统受体的体外结合实验。Phytomedicine。2004;11(7-8):633-638。.https://doi.org/10.1016/j.phymed.2004.03.005
[B305] Brazier NC, Levine MA. 常用西药与草药间的相互作用:医疗保健专业人士名录。Am J Ther。2003;10(3):163-169。.https://doi.org/10.1097/00045391-200305000-00003
[B306] De Groot AC, Schmidt E. 精油:接触性过敏和化学成分. CRC出版社;2016年.https://www.routledge.com/Essential-Oils-Contact-Allergy-and-Chemical-Composition/de-Groot-Schmidt/p/book/9781498702508
[B307] Shellie R, Marriott P, Huie C. 亚洲和美洲人参的二维气相色谱 (GC × GC) 和 GC × GC-四极杆质谱分析。J Sep Sci。2003;26(12-13):1185-1192。.https://doi.org/10.1002/jssc.200301521
[B308] Kubeczka KH. 精油研究的历史和来源。载: Baser KHC, Buchbauer G, 主编. 精油手册:科学、技术与应用. CRC出版社;2010:3-38.https://www.routledge.com/Handbook-of-Essential-Oils-Science-Technology-and-Applications/Baser-Buchbauer/p/book/9781032046419
[B309] Cavanagh HM, Wilkinson JM. 薰衣草精油:一篇综述。澳大利亚感染控制杂志。2005;10(1):35-37。.https://doi.org/10.1071/HI05035
[B310] 梁爱云, 福斯特 S. 食用、药物和化妆品中常用天然成分百科全书. 第2版. 威利出版社; 1996.https://www.wiley.com/en-us/Encyclopedia+of+Common+Natural+Ingredients+Used+in+Food%2C+Drugs%2C+and+Cosmetics%2C+2nd+Edition-p-9780471508267
[B311] Lis-Balchin M. 精油芳香疗法. 载于: Lis-Balchin M, 主编. 芳香疗法科学:医疗专业人员指南. 医药出版社; 2006:49-115.https://www.pharmpress.com/product/9780853696230/aromatherapy-science
[B312] 威廉姆斯 TI. 对芳香疗法的一瞥. Perspect Psychiatr Care. 2006;42(2):107-108.https://doi.org/10.1111/j.1744-6163.2006.00056.x
[B313] Brondino N, Fusar-Poli L, Rocchetti M, Provenzani U, Barale F, Politi P. 孤独症谱系障碍的补充与替代疗法。循证补充替代医学。2015;2015:258589。.https://doi.org/10.1155/2015/258589
[B314] Bent S, Bertoglio K, Hendren RL. Omega-3脂肪酸对孤独症谱系障碍的疗效:一项系统评价。J Autism Dev Disord。2009;39(8):1145-1154。.https://doi.org/10.1007/s10803-009-0724-5
[B315] Masi A, Quintana DS, Glozier N, Lloyd AR, Hickie IB, Guastella AJ. 自闭症谱系障碍中的细胞因子异常:系统评价与荟萃分析。Mol Psychiatry。2015;20(4):440-446。.https://doi.org/10.1038/mp.2014.59
[B316] Veenstra-VanderWeele J, Blakely RD. 孤独症中的网络化:在寻找新疗法的过程中利用遗传学、生物标志物和模型系统研究的发现。《神经精神药理学杂志》。2012;37(1):196-212。.https://doi.org/10.1038/npp.2011.185
[B317] Yamasue H, Domes G. 催产素与自闭症谱系障碍. Curr Top Behav Neurosci. 2018;35:449-465.https://doi.org/10.1007/7854_2017_24
[B318] Petra AI, Panagiotidou S, Hatziagelaki E, Stewart JM, Conti P, Theoharides TC. 肠道-微生物-大脑轴及其对怀疑免疫失调的神经精神疾病的影响。Clin Ther. 2015;37(5):984-995。.https://doi.org/10.1016/j.clinthera.2015.04.002
[B319] Mayer EA, Tillisch K, Gupta A. 肠道/大脑轴与微生物群。《临床研究杂志》。2015;125(3):926-938。.https://doi.org/10.1172/JCI76304
[B320] Button KS, Ioannidis JP, Mokrysz C, et al. 供电故障:为什么小样本量会削弱神经科学的可靠性。Nat Rev Neurosci。2013;14(5):365-376。.https://doi.org/10.1038/nrn3475
[B321] 希金斯 JP, 汤普森 SG, 迪克斯 JJ, 奥特曼 DG. Meta分析中不一致性的测量. BMJ. 2003;327(7414):557-560.https://doi.org/10.1136/bmj.327.7414.557
[B322] Rossignol DA. 自闭症谱系障碍的新颖和新兴疗法:系统评价。Ann Clin Psychiatry。2009;21(4):213-236。.https://pmc.ncbi.nlm.nih.gov/articles/PMC3859980/
[B323] Moher D, Liberati A, Tetzlaff J, Altman DG; PRISMA Group. 系统评价和荟萃分析的首选报告项目:PRISMA声明。PLoS Med. 2009;6(7):e1000097。.https://doi.org/10.1371/journal.pmed.1000097
[B324] Ioannidis JP, Patsopoulos NA, Rothstein HR. Meta分析在森林图中的回避理由或借口. BMJ. 2008;336(7658):1413-1415.https://doi.org/10.1136/bmj.a117
[B325] Loth E、Charman T、Mason L 等人。欧盟-AIMS 纵向欧洲自闭症项目 (LEAP):识别和验证自闭症谱系障碍分层生物标志物的设计和方法。Mol Autism。2017;8:24。.https://doi.org/10.1186/s13229-017-0146-8
[B326] Geschwind DH, Flint J. 精神疾病的遗传学和基因组学. 科学. 2015;349(6255):1489-1494.https://doi.org/10.1126/science.aaa8954
[B327] Easterbrook PJ, Berlin JA, Gopalan R, Matthews DR. Publication bias in clinical research. Lancet. 1991;337(8746):867-872.https://doi.org/10.1016/0140-6736(91 )90201-y
[B328] Nestler EJ, Hyman SE. 神经精神疾病的动物模型. Nat Neurosci. 2010;13(10):1161-1169.https://doi.org/10.1038/nn.2647
[B329] Buchbauer G, Jirovetz L, Jäger W, Plank C, Dietrich H. 具有吸入镇静作用的香料化合物和精油。J Pharm Sci. 1993;82(6):660-664.https://doi.org/10.1002/jps.2600820623
[B330] Cooke B, Ernst E. 芳香疗法:一项系统性评价。英国全科医师杂志。2000;50(455):493-496。.https://pubmed.ncbi.nlm.nih.gov/10962794/
【B331】Halm MA. 精油在危重病人症状管理中的应用。《美国重症监护杂志》. 2008;17(2):160-163.https://www.researchgate.net/publication/5541002_Essential_Oils_for_Management_of_Symptoms_in_Critically_Ill_Patients
[B332] Hammer KA, Carson CF, Riley TV. 精油及其他植物提取物的抗菌活性。《应用微生物学杂志》1999;86(6):985-990。.https://doi.org/10.1046/j.1365-2672.1999.00780.x
[B333] Schulz KF, Altman DG, Moher D; CONSORT Group. CONSORT 2010声明:平行分组随机试验报告更新指南。BMJ。2010;340:c332。.https://doi.org/10.1136/bmj.c332
[B334] Lombardo MV, Lai MC, Baron-Cohen S. 自闭症谱系异质性分解的大数据方法。Mol Psychiatry。2019;24(10):1435-1450。.https://doi.org/10.1038/s41380-018-0321-0
[B335] Insel TR. NIMH 研究领域标准(RDoC)项目:精神病学的精准医疗。《美国精神病学杂志》。2014;171(4):395-397。.https://doi.org/10.1176/appi.ajp.2014.14020138
[B336] Jeste SS,Geschwind DH。神经发育障碍的临床试验:治疗前沿。Sci Transl Med。2016;8(321):321fs1。.https://doi.org/10.1126/scitranslmed.aad9874
[B337] Kovar KA, Gropper B, Friess D, Ammon HP. 迷迭香油吸入和口服后小鼠的1,8-桉叶素血药浓度和运动活动。Planta Med. 1987;53(4):315-318。.https://doi.org/10.1055/s-2006-962725
[B338] Rossignol DA, Frye RE. 自闭症谱系障碍生理异常研究趋势的综述:免疫失调、炎症、氧化应激、线粒体功能障碍和环境毒素暴露。《分子精神病学》。2012;17(4):389-401。.https://doi.org/10.1038/mp.2011.165
[B339] Stang A. 对用于荟萃分析中对非随机研究质量进行评估的Newcastle-Ottawa量表的批判性评价。Eur J Epidemiol. 2010;25(9):603-605。.https://doi.org/10.1007/s10654-010-9491-z
[B340] Herz RS. 香薰疗法:对情绪、生理和行为的嗅觉影响的科学分析。国际神经科学杂志。2009;119(2):263-290。.https://doi.org/10.1080/00207450802333953
[B341] Myers SM, Johnson CP; 美国儿科学会残疾儿童委员会。自闭症谱系障碍儿童的管理。《儿科学》杂志。2007;120(5):1162-1182。.https://doi.org/10.1542/peds.2007-2362
[B342] Volkmar F, Siegel M, Woodbury-Smith M, King B, McCracken J, State M;美国儿童和青少年精神病学学会(AACAP)质量问题委员会(CQI)。用于评估和治疗自闭症谱系障碍儿童和青少年的实践参数。J Am Acad Child Adolesc Psychiatry。2014;53(2):237-257。.https://doi.org/10.1016/j.jaac.2013.10.013
[B343] Green VA, Pituch KA, Itchon J, Choi A, O’Reilly M, Sigafoos J. 互联网调查自闭症儿童家长使用的疗法。《残疾研究与发展杂志》。2006;27(1):70-84。.https://doi.org/10.1016/j.ridd.2004.12.002
[B344] Bilia AR, Guccione C, Isacchi B, Righeschi C, Firenzuoli F, Bergonzi MC. 负载于纳米系统的精油:成功治疗策略的新发展。Evid Based Complement Alternat Med. 2014;2014:651593.https://doi.org/10.1155/2014/651593
[B345] Woolf AD. 精油中毒. J Toxicol Clin Toxicol. 1999;37(6):721-727.https://doi.org/10.1081/clt-100102450
[B346] Kasper S, Gastpar M, Müller WE, 等. Silexan,一种口服薰衣草油制剂,在亚阈值焦虑障碍中的疗效与安全性——来自临床试验的证据. Wien Med Wochenschr. 2010;160(21-22):547-556.https://doi.org/10.1007/s10354-010-0845-7
生姜补充剂通过调节产前丙戊酸暴露小鼠的AKT/GSK3β信号通路来缓解自闭症行为。. 医学杂志. https://doi.org/10.1039/d5fo04364f
B348. 波斯医药产品Hiwa糖浆对自闭症谱系障碍患儿自闭症症状的影响:一项随机双盲临床试验。. 全球医学杂志. https://doi.org/10.31661/gmj.v13i.3553
B349. McGuinness, F., Oswald, I. W. H., Koby, K. A. 等人 (2023)。大麻二酚和受大麻启发的萜烯混合物在自闭症谱系障碍的 BTBR 小鼠模型中具有急性亲社会作用。. 神经科学前沿. https://doi.org/10.3389/fnins.2023.1185737
ADHD 词汇表
注意缺陷/多动障碍
注意力缺陷多动障碍.
辅助治疗
辅助治疗,除主要治疗外使用,以增强其疗效或减轻副作用。.
抗焦虑药
舒缓焦虑,减轻焦虑。.
血脑屏障
血脑屏障,控制物质进入大脑的选择性屏障。.
CB2受体
大麻素受体 2 型,内源性大麻素系统的一部分,主要存在于免疫细胞上;激活具有抗炎作用。.
化学类型
植物的化学变体,其精油成分不同(例如,迷迭香化学型桉叶油素与樟脑)。.
DAT(多巴胺转运体)
将多巴胺从突触间隙转运回神经元的蛋白质;哌甲酯的主要靶点。.
DAT-占领
多巴胺转运体被药物阻断的百分比。.
多巴胺
神经递质,对动机、奖励、运动和注意力很重要;ADHD患者通常含量偏低。.
ED₅₀
达到最大效应% 的 50% 的剂量。.
GABA(γ-氨基丁酸)
大脑中最主要的抑制性神经递质;降低神经元兴奋性。.
GABA-A受体
GABA离子通道受体;激活导致氯离子内流和神经元超极化(抑制)。.
气相色谱-质谱联用
鉴别和定量精油中化学成分的分析方法。.
IC₅₀
浓度,达到目标(例如,受体、酶)最大抑制的 50%。.
边缘系统
处理情绪、记忆和动机的大脑区域;包括杏仁核、海马体和其他结构。.
芳樟醇
单萜醇,薰衣草的主要活性成分;具有抗焦虑、镇静作用,并调节神经递质系统。.
薄荷醇
薄荷单萜烯醇;激活TRPM8冷受体,具有清爽和提高注意力的作用。.
盐酸哌甲酯
兴奋剂,抑制多巴胺和去甲肾上腺素再摄取;ADHD 的一线治疗。.
微透析
活体大脑中细胞外神经递质浓度测量技术。.
单胺
一组神经递质,包括多巴胺、去甲肾上腺素和血清素。.
去甲肾上腺素转运体
将去甲肾上腺素从突触间隙运输回神经元的蛋白质;也是哌甲酯的作用靶点。.
神经炎症
大脑中的炎症过程,通常由激活的小胶质细胞和促炎细胞因子介导。.
神经递质
在神经细胞之间传递信号的化学信使(例如,多巴胺、血清素、GABA)。.
NF-κB(核因子κB)
转录因子,激活炎症基因;抑制具有抗炎作用。.
NMDA受体
谷氨酸受体,对突触可塑性和学习很重要;过度激活可能具有神经毒性。.
去甲肾上腺素
神经递质,对清醒、注意力、压力反应很重要;ADHD 患者通常有所缺乏。.
占领
参见 DAT-占有率;受体或转运蛋白的占有百分比。.
嗅觉
关于嗅觉。.
正电子发射断层显像
利用放射性标记物质来可视化大脑代谢过程和受体结合的成像技术。.
药物动力学
学习药物在体内的过程(吸收、分布、代谢、排泄)。.
药效学
关于药物在体内作用(作用机制、效果)的教学。.
前额叶皮层
额叶后区,对执行功能(计划、冲动控制、工作记忆)很重要;多动症时常活动不足。.
受体
结合特异性信号分子(神经递质、激素)并由此引发细胞反应的细胞内或细胞上(膜上)的蛋白质。.
血清素转运体
将血清素从突触间隙泵回神经元的蛋白质;抗抑郁药(SSRIs)的作用靶点。.
羟色胺
神经递质,对情绪、睡眠、食欲和冲动控制很重要。.
单光子发射计算机断层扫描
一种成像技术,类似于 PET,使用放射性示踪剂来描绘大脑功能。.
纹状体
基底神经节的脑区,富含多巴胺转运体;对运动、动机和奖励很重要;哌甲酯的主要靶区域。.
突触
两个神经元之间的连接点,在此处释放神经递质。.
突触间隙
突触处两个神经元之间的狭窄空间,神经递质在此释放。.
瞬时受体电位通道
对不同刺激(温度、化学物质)有反应的离子通道家族;例如,TRPM8(冷/薄荷醇),TRPV1(热/辣椒素)。.
文丘里效应
一种物理原理,通过在狭窄处压强降低,将液体或油吸入并雾化;许多扩散器的基础。.
β-石竹烯
黑胡椒和大麻中的倍半萜;具有抗炎特性的选择性CB2受体激动剂。.
1,8-桉叶素 (桉叶油精)
来自迷迭香和桉树的单萜氧化物;具有祛痰、消炎和促进认知作用。.
5-HT1A受体
血清素受体亚型;激活具有抗焦虑和抗抑郁作用。.
报告结束
本报告于2026年4月27日根据对87项关于哌甲酯脑浓度的研究和159项关于ADHD背景下精油的文献进行的系统性分析而撰写。所有陈述均有原始文献支持。.
ASD 词汇表
1,8-桉叶素 (桉叶油精)
单萜氧化物,桉树油的主要成分(70–90 %),通过GABA-A受体调节显示出抗焦虑作用和抗炎特性;因喉痉挛风险,禁忌用于3岁以下儿童。.
α-蒎烯
松树、迷迭香和**大麻**中的双环单萜;显示GABA能调节、抗焦虑和抗炎作用;临床前证据表明其具有神经保护作用。.
杏仁核
杏仁核样的脑部结构位于内侧颞叶,是情绪处理、恐惧条件反射和社会感知的中枢;在自闭症谱系障碍(ASD)患者的幼儿时期会增大,在面部处理时活动性降低。.
抗氧化剂
一种通过中和活性氧 (ROS) 来减少氧化应激的分子;例如:谷胱甘肽、超氧化物歧化酶、过氧化氢酶、维生素 C、维生素 E;在 ASD 中抗氧化能力降低。.
应用行为分析
循证的自闭症行为干预,基于学习原则(强化、提示、塑形);改善沟通、社交技能和适应性行为。.
阿立哌唑
非典型抗精神病药物,部分多巴胺 D2 受体激动剂;FDA 批准用于自闭症谱系障碍(ASD)的易怒;副作用:体重增加、静坐不能、代谢综合征;通过 CYP2D6 和 CYP3A4 代谢。.
星形胶质细胞
中枢神经系统中的星形胶质细胞;通过 EAAT1/2 转运体调节谷氨酸稳态、K+ 缓冲、血脑屏障和神经元代谢支持;在自闭症谱系障碍 (ASD) 中表现为星形胶质细胞增生(GFAP 表达升高)。.
β-茶碱(BCP)
胡椒、丁香、大麻中的倍半萜类碳氢化合物;选择性 CB2 受体激动剂,无精神活性作用;可减轻神经炎症、小胶质细胞活化和氧化应激。.
苯二氮䓬结合位点
GABA-A受体上的变构调节位点;苯二氮䓬类(地西泮、劳拉西泮)在此结合并增强GABA能抑制;芳樟醇和1,8-桉叶油精与此位点相互作用(氟马西尼敏感)。.
佛手柑(香柠檬)
柑橘类水果,其精油含有芳樟醇、乙酸芳樟酯、柠檬烯;一项针对ASD儿童的RCT显示没有显著的抗焦虑效果;含有佛手柑内酯(CYP3A4抑制剂),有不含佛手柑内酯的变种(FCF)。.
乳香酸
来自乳香的五环三萜酸;激活 Nrf2/HO-1 抗氧化通路,减少神经炎症(小胶质细胞、星形胶质细胞、TNF-α、IL-6);用于改善 VPA 模型中 ASD 样行为的临床前证据。.
布美他尼
袢利尿剂, NKCC1 协同转运蛋白抑制剂; 纠正 ASD 小鼠模型中延迟的 GABA 开关; 临床研究表明 ASD 儿童社交沟通能力有适度改善, 但存在方法学上的局限性。.
樟脑
樟脑(Cinnamomum camphora)和迷迭香(樟脑化学型)中的萜类酮;降低癫痫阈值,可能诱发癫痫发作;癫痫患者和患有合并癫痫的ASD患者禁用。.
依兰
热带树木,其精油含有芳樟醇、香叶醇、β-石竹烯;临床前研究表明,通过调节血清素和多巴胺代谢,可改善VPA大鼠的自闭症样行为。.
CB2 受体(2 型大麻素受体)
G蛋白偶联受体,主要在免疫细胞(小胶质细胞、外周免疫细胞)上表达;激活可减少细胞因子释放、小胶质细胞活化和神经炎症;β-石竹烯是选择性CB2激动剂。.
雪松醇
雪松油中的倍半萜烯醇;传统上认为具有镇静作用,但缺乏关于GABA能机制或在自闭症谱系障碍(ASD)中临床疗效的科学证据。.
拷贝数变异 (CNVs)
基因组缺失或重复>1 kb;在ASD中常见(15q11-13重复,16p11.2缺失,22q13缺失/Phelan-McDermid综合征);常累及突触基因(SHANK3,NLGN4)。.
细胞色素P450 (CYP)
催化药物代谢的酶家族;CYP3A4 和 CYP2D6 代谢利培酮和阿立哌唑;佛手柑内酯(在佛手柑中)抑制 CYP3A4,可能升高药物水平。.
多巴胺 (DA)
儿茶酚胺类神经递质;介导奖赏处理、动机、运动控制;在自闭症谱系障碍中,社会奖赏相关的纹状体激活减少,多巴胺转运体密度改变;依兰依兰在临床前模型中调节多巴胺代谢。.
精神疾病诊断与统计手册,第五版
美国精神病学协会的分类系统;通过两个主要领域定义自闭症谱系障碍:(1) 社交沟通/互动方面的持续性缺陷;(2) 受限、重复的行为模式。.
菌群失调
肠道菌群失调;孤独症谱系障碍中微生物多样性降低,厚壁菌门/拟杆菌门比例增加,双歧杆菌和普雷沃菌减少;与症状严重程度相关。.
兴奋-抑制失衡
兴奋性(谷氨酸能)和抑制性(GABA能)神经传递失衡;自闭症谱系障碍的核心病理生理学假说;导致过度兴奋、感觉过滤障碍和社交信息处理受损。.
电针
电针;临床前研究表明,通过迷走神经依赖性机制和微生物组调节,改善 VPA 小鼠的自闭症样行为。.
癫痫
Neurologische Erkrankung mit rekurrenten Anfällen; 10–30 % der ASD-Patienten haben komorbide Epilepsie; campherhaltige ätherische Öle (Kampfer, Rosmarin-Campher-Chemotyp, Salbei) sind kontraindiziert.
粪便微生物移植 (FMT)
粪菌移植,受体接受来自健康捐赠者的粪菌;临床前研究表明,在ASD小鼠模型中具有类ASD行为的改善;在ASD儿童开放性临床试验中显示了胃肠道症状和行为的改善。.
氟马西尼
苯二氮䓬受體拮抗劑,結合到GABA-A受體的苯二氮䓬結合位點;阻斷芳樟醇和1,8-桉樹腦的抗焦慮作用,證實了它們與該位點的相互作用。.
GABA(γ-氨基丁酸)
中枢神经系统主要的抑制性神经递质;与GABA-A(离子型)和GABA-B(代谢型)受体结合;在ASD中GABA能功能减退、GABA水平改变、发育过程中GABA开关受损。.
GABA-A受体
配体门控氯离子通道;GABA激活导致氯离子内流和超极化(抑制);含有苯二氮䓬结合位点;芳樟醇、1,8-桉树脑和α-蒎烯可调节GABA-A受体。.
GABA-开关
兴奋性到抑制性GABA作用的神经发育转化;由NKCC1到KCC2共转运蛋白表达的转变介导;在ASD中延迟;催产素和布美他尼可以使GABA转换正常化。.
谷氨酸脱羧酶65/67
将谷氨酸转化为GABA的酶;在ASD中,前额叶皮层和小脑的GAD65/67表达减少,导致GABA合成减少。.
气相色谱-质谱联用(GC/MS)
用于鉴定和定量挥发性成分的分析技术;精油化学表征的金标准;能够检测掺假、污染物和批次间差异。.
谷氨酸 (Glu)
中枢神经系统主要的兴奋性神经递质;与离子型(NMDA、AMPA、海藻酸盐)和代谢型(mGluR1-8)受体结合;自闭症谱系障碍(ASD)中谷氨酸水平改变、谷氨酸能突触基因(GRIN2B、GRIA1)受损、谷氨酸能传递过度。.
谷胱甘肽
三肽(γ-谷氨酰-半胱氨酰-甘氨酸),主要的细胞内抗氧化剂;自闭症谱系障碍(ASD)中谷胱甘肽(GSH)水平降低,氧化型谷胱甘肽(GSSG)升高,GSH/GSSG比率降低;GSH合成酶(GCLC、GSS)的基因变异与ASD相关。.
肠脑轴
胃肠道和中枢神经系统之间的双向通讯,通过神经(迷走神经)、内分泌(HPA轴)、免疫和代谢途径;在自闭症谱系障碍中,肠道菌群失调,神经递质前体产生改变,肠脑信号通路紊乱。.
血红素加氧酶-1 (HO-1)
一种酶,它将血红素分解为胆绿素(抗氧化)、一氧化碳(抗炎)和铁;Nrf2 调控;乳香酸能增加 HO-1 的表达;HO-1 诱导能抵抗氧化应激和神经炎症。.
海马体
内侧颞叶的脑部结构;介导陈述性记忆、空间导航、情境处理;在自闭症谱系障碍儿童期增大,连接改变,情境恐惧条件化受损。.
高血清素血症
Erhöhte periphere Serotonin-Spiegel; bei 25–50 % der ASD-Patienten nachgewiesen; Beziehung zu zentralen 5-HT-Spiegeln unklar; möglicherweise durch gestörte Plättchen-Serotonin-Aufnahme oder erhöhte periphere Synthese.
白细胞介素-6 (IL-6)
促炎细胞因子;在孤独症患者的血浆和脑脊液中IL-6水平升高;母体免疫激活伴IL-6产生在小鼠模型中产生孤独症样表型;IL-6调节突触传递和发育。.
小脑
脑干结构;传统上与运动控制有关,但也参与认知过程、感觉预测和社会学习;在自闭症谱系障碍中,浦肯野细胞丢失、小脑蚓部体积减小、小脑皮质连接受损。.
薰衣草 (狭叶薰衣草)
芳香植物,精油含% 25-45% 的芳樟醇和乙酸芳樟酯;小型试点研究报告称,自闭症谱系障碍儿童的 ABA 依从性和注意力分配有所改善;临床前证据表明,其具有 GABA 能调节、5-羟色胺能调节和催产素能激活作用。.
芳樟醇
单萜烯醇,薰衣草、芫荽、佛手柑的主要成分;调节GABA-A受体在苯二氮䓬结合位点(氟马西尼敏感),影响血清素水平,增加催产素神经元细胞内钙,表现出抗炎作用。.
乙酸芳樟酯
一种由芳樟醇和乙酸形成的酯;薰衣草(25–45 %)和佛手柑的主要成分;有助于抗焦虑和镇静作用;水解为芳樟醇。.
长期增强 (LTP) / 长期抑制 (LTD)
突触可塑性的细胞机制;长期增强(LTP):高频刺激后突触传递的持续增强;长期抑制(LTD):低频刺激后突触传递的持续减弱;自闭症谱系障碍(ASD)中,多脑区LTP/LTD平衡失调。.
磁共振波谱
一种用于体内定量代谢物(谷氨酸、GABA、N-乙酰天冬氨酸、肌酸)的非侵入性成像技术;在 ASD 中,MRS 显示谷氨酸和 GABA 浓度存在区域性改变。.
母体免疫激活
一种模型,在该模型中,孕期动物接受免疫刺激剂(Poly(I:C)、LPS)治疗;后代表现出类似 ASD 的行为,促炎细胞因子增加,突触发育受损;模拟了 ASD 的环境风险因素。.
褪黑素
一种由松果体产生的激素;调节昼夜节律和睡眠;自闭症谱系障碍患儿常有睡眠障碍;褪黑素补充剂可改善入睡潜伏期和睡眠持续时间;荟萃分析显示具有中等疗效。.
小胶质细胞
中枢神经系统常驻免疫细胞;调节突触修剪、神经元发育、免疫监视;自闭症谱系障碍中,微生物胶质细胞(Iba1表达升高,变形虫形态)在中枢前额叶皮层和小脑中被激活;释放促炎细胞因子和活性氧。.
线粒体功能障碍
Beeinträchtigte mitochondriale Funktion (reduzierte Atmungsketten-Aktivität, erhöhtes Laktat, reduzierte ATP-Synthese); bei 5–30 % der ASD-Patienten; führt zu erhöhter ROS-Produktion, oxidativem Stress, gestörter Calcium-Homöostase.
神经细胞黏附分子(NLGN)
突触后细胞粘附分子(NLGN1-4);与突触前ネーレキシン相互作用;调节突触分化、成熟和功能;NLGN3和NLGN4X中的突变见于ASD患者;NLGN3-R451C小鼠表现出抑制性传递增加和ASD样表型。.
神经细胞粘附分子 (NRXN)
突触前细胞粘附分子(NRXN1-3);与突触后神经粘素相互作用;调节突触组织;NRXN1在ASD患者中的突变;NRXN1缺失与ASD、精神分裂症和智力障碍相关。.
神经炎症
中枢神经系统炎症反应;以小胶质细胞和星形胶质细胞激活、促炎细胞因子(TNF-α、IL-6、IL-1β)升高、血脑屏障受损为特征;在ASD中,多脑区存在慢性神经炎症。.
NF-κB(核因子κB)
一种转录因子,可调节促炎基因(TNF-α、IL-6、IL-1β、COX-2);自闭症谱系障碍中 NF-κB 激活增加;β-石竹烯和乳香酸可抑制 NF-κB 信号通路。.
NMDA受体 (N-甲基-D-天冬氨酸受体)
离子型谷氨酸受体,钙离子通透性通道;突触可塑性(LTP/LTD)、学习和记忆的核心;ASD中NMDA受体亚基表达改变(GRIN2A,GRIN2B),SHANK2基因敲除小鼠中NMDA受体功能受损。.
Nrf2 (核因子红系2相关因子2)
转录因子,抗氧化反应的 master 调控因子;诱导 HO-1、NQO1、谷胱甘肽 S-转移酶和其他抗氧化酶的表达;乳香酸活化 Nrf2;Nrf2 激活剂(萝卜硫素)在小型研究中显示出对 ASD 的改善。.
氧化应激
ROS生成与抗氧化防御之间的不平衡;ASD中脂质过氧化、蛋白质羰基化、DNA氧化增加,抗氧化能力(GSH、SOD、过氧化氢酶)降低;导致神经损伤和功能障碍。.
催产素(OT)
神经肽,由下丘脑产生;在社会纽带、信任、社会认知、压力调节中起核心作用;ASD 患者血浆 OT 水平降低,OXTR 表达改变;鼻腔内 OT 结果不一;OT 在临床前模型中调节 GABA 能发育。.
पहाडी परवलय-धनात्मक मध्यवर्तीन्यूरॉन
GABA能中间神经元亚型,表达钙结合蛋白视黄醛结合蛋白;介导快速、同步抑制;在ASD患者中,前额叶皮层视黄醛结合蛋白中间神经元密度和功能降低。.
费兰-麦克德米德综合征
22q13缺失综合征,包含SHANK3基因;特征为自闭症谱系障碍、智力障碍、语言发育迟缓、肌张力减退;表明SHANK3在自闭症谱系障碍发病机制中的因果作用。.
前额叶皮层
额叶脑区;介导执行功能、工作记忆、认知灵活性、社会认知、心智理论;在自闭症谱系障碍(ASD)患者的社会认知任务中,额叶脑区(PFC)活动减弱,PFC连接性受损,GABA能缺陷。.
丙酸酯
短链脂肪酸,由肠道微生物群产生;在自闭症谱系障碍中丙酸水平可能升高;在啮齿动物模型中暴露于丙酸可诱导自闭症谱系障碍样行为、神经炎症、氧化应激。.
活性氧 (ROS)
高反应性分子(超氧化物、过氧化氢、羟基自由基);线粒体呼吸的副产物;过量浓度会引起活性氧对脂质、蛋白质、DNA的氧化损伤;自闭症谱系障碍中活性氧产生增加。.
瑞斯喜
非典型抗精神病药物,多巴胺 D2 和血清素 5-HT2A 受体拮抗剂;FDA 批准用于自闭症谱系障碍(ASD)的易怒症状;副作用:体重增加、高催乳素血症、锥体外系症状、代谢综合征;通过 CYP2D6 代谢。.
血清素(5-HT,5-羟色胺)
单胺类神经递质;介导情绪、焦虑、睡眠、食欲、社交认知;在自闭症谱系障碍中,有高血清素血症(外周)、中枢血清素合成减少、5-HTT 和 TPH2 的基因变异;芳樟醇和依兰依兰可调节血清素能系统。.
SHANK蛋白(SH3和多个ankyrin重复结构域)
突触后支架蛋白(SHANK1-3);组装谷氨酸受体、肌动蛋白细胞骨架、信号分子;SHANK2和SHANK3的突变与ASD有关;SHANK3基因敲除小鼠表现出棘突密度降低、突触传递受损、类ASD行为。.
纹状体
基底神经节的皮质下结构(尾状核、壳核、伏隔核);介导奖赏处理、习惯学习、运动控制;自闭症谱系障碍(ASD)中表现为纹状体增大、社会奖赏激活减弱、皮质-纹状体连接障碍。.
突触修剪
发育生物学过程,其中过剩的突触被消除;通过小胶质细胞、补体系统介导;在自闭症谱系障碍中,修剪可能受到干扰(过多或过少),导致突触密度和连接性改变。.
萜烯
一类由异戊二烯单元(C5H8)构成的天然产物;精油的主要成分;单萜(C10):芳樟醇、1,8-桉叶素、α-蒎烯;倍半萜(C15):β-石竹烯、雪松醇;三萜(C30):乳香酸。.
肿瘤坏死因子-α
促炎细胞因子;在自闭症谱系障碍中,血浆、脑脊液、死后脑组织中的 TNF-α 水平升高;母体免疫激活伴随 TNF-α 升高可产生类似自闭症谱系障碍的表型;TNF-α 调节突触传递,降低 GABA 能抑制。.
转运蛋白 (TSPO)
线粒体蛋白,在激活的小胶质细胞中上调;PET 配体(例如 [11C]-PBR28)与 TSPO 结合,可实现小胶质细胞激活的体内成像;在 ASD 中,多个脑区的 TSPO 结合增加。.
色氨酸
必需氨基酸,血清素和褪黑素的前体;在自闭症谱系障碍中可能存在色氨酸代谢紊乱;肠道微生物群通过犬尿氨酸和血清素通路影响色氨酸的可用性和代谢。.
丙戊酸
抗癫痫药;产前VPA暴露是人类ASD的危险因素;VPA诱导的ASD小鼠模型:孕鼠接受VPA,后代表现出ASD样行为、神经炎症、氧化应激;广泛的临床前模型。.
文丘里效应
物理原理(伯努利效应):当流体流经收缩部位时,其压力会降低;在 Organik Aromas Nebulizer 3.0 中,压缩空气通过喷嘴产生负压,将精油吸入并雾化成细小液滴(冷雾化)。.
香根草 (Vetiveria zizanioides)
热带草本植物,精油含有香根草醇、留兰香醇、α-香根酮;有传闻称对注意力和多动有影响,但在自闭症谱系障碍中缺乏对照研究;作用机制的科学证据不足。.
乳香(乳香)
乳香属植物的树脂;精油和提取物含有乳香酸;临床前证据表明具有抗神经炎症、抗氧化(Nrf2/HO-1)和神经保护作用;在自闭症谱系障碍(ASD)方面未发现临床研究。.
雪松木(大西洋雪松, 弗吉尼亚刺柏)
精油含有雪松醇、α-雪松烯、侧柏酮;传统上用于镇静和缓解焦虑,但缺乏关于 GABA 能机制或在 ASD 中的临床疗效的科学证据。.
昼夜节律
内源性~24小时节律,调节睡眠-觉醒周期、激素分泌、体温;ASD患者常有昼夜节律紊乱、睡眠障碍;褪黑素可调节昼夜节律并改善ASD患者的睡眠。.