Innehållsförteckning
Lesezeit 187 Minuten
Uppdaterad – 30 april 2026
En jämförande studie av standardbehandling för ADHD/ADD och aromaterapi med terapeutiskt godkända eteriska oljor, batchvis testade med GC/MS-analyser. (Status 04.2026).
ADHD/ADD och eteriska oljor – förklarat på ett begripligt sätt
En lättförståelig sammanfattning av den vetenskapliga rapporten för alla som vill lära sig mer om naturligt stöd vid ADHD
Vad är ADHD/ADD?
ADHD står för ADHD, ADS för varianten utan hyperaktivitet (uppmärksamhetsunderskottstörning). Båda är inte påhitt eller uppfostringsfel, de är verkliga neurologiska tillstånd där hjärnan är annorlunda kopplad.
Bilden bakom: Föreställ dig hjärnan som en radio. Hos en ADHD-hjärna hoppar sändaren ständigt mellan frekvenser, ibland för högt, ibland för lågt, sällan på rätt kanal. Det gör det svårt att koncentrera sig, sitta stilla eller kontrollera impulser.
Vad händer i hjärnan vid ADHD?
Kärnproblemet ligger i två signalämnen:
Dopamin – den “belönings- och motivationsbudbäraren”
Vid ADHD “städades” dopamin för snabbt upp igen (genom dopamintransportören DAT). Detta leder till bristande känsla av belöning och motivation för vardagliga uppgifter. Hjärnan söker ständigt stimulans, därav impulsivitet och risktagande.
Noradrenalin – “uppmärksamhets- och fokusbudbäraren”
För lite noradrenalin i prefrontala cortex (hjärnans “chefområde”) – Leder till koncentrationsproblem, glömska, svårigheter att planera
Vilka hjärnregioner påverkas?
- Prefrontala cortex
Ansvarig för planering, impulskontroll, koncentration – underaktiv vid ADHD
- Striatum (Basala ganglierna)
Belöningscentrum – mindre känsligt vid ADHD
- Lillhjärnan
Timing och koordination – lätt förändrat vid ADHD
Hur vanligt är ADHD?
Hos barn och ungdomar är det den vanligaste psykiatriska sjukdomen med 5 till 7 %, hos vuxna 2,5 till 4 %, många vuxna vet inte att de har ADHD.
Hur yttrar sig ADHD?

Hur behandlas ADHD vanligtvis?
Huvudläkemedlet – metylfenidat (Ritalin, Concerta, Medikinet)
Metylfenidat (MPH) är det vanligaste förskrivna ADHD-läkemedlet och förstahandsbehandlingen.
Hur fungerar det?
Metylfenidat blockerar dopamintransportören (DAT), “städ-pumpen” för dopamin. När färre dopaminmolekyler tas upp, stannar mer kvar mellan nervcellerna, och hjärnan kan fokusera bättre.
Siffrorna bakom
En dos på 20 mg blockerar ungefär. 54 % dopaminrecirkuleringspumpen i hjärnan, 40 mg blockerar ungefär. 72 %.
Effekten inträder efter 30–90 minuter och varar beroende på formulering 4–12 timmar.
Hur effektivt är det?
Med 70-80 % Patienten svarar kliniskt bra på metylfenidat: förbättringar av uppmärksamhet, impulsivitet och skolprestationer är väl dokumenterade.
Formuleringar
- Kortverkande (t.ex. Ritalin): 4–6 timmar, måste tas flera gånger om dagen
- Retarderad (t.ex. Concerta, Medikinet retard): 8–12 timmar, en gång dagligen
Biverkningar
– Aptitnedsättning (vanligaste biverkningen)
– Sömnstörningar (om det tas för sent)
– Huvudvärk, magbesvär (oftast i början)
– Lätt ökning av puls och blodtryck
– Humörsvängningar, irritabilitet (sällan)
Ytterligare läkemedel
Atomoxetin (Strattera)
– Ingen stimulans, hämmar noradrenalintransportören (NET)
– Verkar även på ADHD utan missbruksrisk
– Tar 4–6 veckor för full effekt
– Lämplig vid samtidig ångeststörning eller ticsyndrom
Amfetamin (i Tyskland mindre vanligt)
– Lisdexamfetamin (Vyvanse): Godkänd i Tyskland sedan 2013 för vuxna
– Verkar starkare och längre än metylfenidat
Guanfacin (Intuniv)
– För barn och ungdomar där stimulantia inte fungerar
– Verkar på noradrenalinreceptorer i prefrontala cortex
Eteriska oljor vid ADHD – hur kan det fungera?
Eteriska oljor kan stödja ADHD-hjärnan på flera sätt:
- Om luktsinnet till hjärnan
Doftmolekyler aktiverar det limbiska systemet (känslor, motivation) och prefrontala cortex (koncentration) – direkt och snabbt - Om signalsubstanssystem
Vissa terpener påverkar dopamin, noradrenalin, serotonin och GABA - Komplement till metylfenidat
Metylfenidat verkar på DAT/NET – eteriska oljor verkar på andra system (GABA-A, CB2, TRPM8, SERT) och är därmed potentiella komplement
Viktig begränsning
Forskningen kring eteriska oljor vid ADHD är fortfarande i sin linda. De flesta studier har gjorts på friska vuxna, inte på patienter med ADHD. Direkta kliniska bevis är fortfarande begränsade.
Vilka eteriska oljor kan hjälpa vid ADHD?
Pepparmynta – koncentrationsoljan nr 1
Pepparmint (Mentha × piperita) har det starkaste beviset för förbättrad uppmärksamhet.
- Vad finns det i?
Mentol (30–50 %)
Menton (10–30 %)
1,8-Cineol (5–10 %) - Mentol kyler huden genom att binda sig till känsligare receptorer i huden, som kallas för TRPM8-receptorer. När mentol binder sig till dessa receptorer uppstår ett kyleffekt som kan kännas långvarig.
Aktiverar TRPM8-receptorer – “kallreceptorer” i nervsystemet, som aktiverar hjärnan
Hemmat enzymet som bryter ner signalsubstansen acetylkolin
Modulerar GABA-A-receptorer (lätt lugnande vid överaktivitet) - Klinisk studie (n=24)
100 µL pepparmyntsolja (inkapslad) förbättrade signifikant kontinuerlig uppmärksamhet i RVIP-testet (Rapid Visual Information Processing) efter 1–3 timmar; minskade trötthet och förbättrade mental aritmetik - Särskilt inslag
Pepparmynta främjar fokuserad Uppmärksamhet (i motsats till lavendel, som snarare sprider uppmärksamheten) - Hur man applicerar
2 droppar på en näsduk, strax före lärande- eller arbetspass; diffusion i studierummet
Rosmarin – för minne och vakenhet
Rosmarin (Rosmarinus officinalis) har sedan antiken kallats “minnesört”, med rätta.
- Vad finns det i?
1,8-Cineol (20–50 %)
Kamfer (10–20 %)
α-Pinen (10–15 %) - Vad gör 1,8-Cineol?
Hämmt enzymet acetylkolinesteras, som bryter ner signalsubstansen (liknande Alzheimers medicin!)
Aktiverar nervsystemet, ökar vakenhet och medvetenhet
Antiinflammatorisk och antioxidant i hjärnan - Klinisk studie (n=144)
Rosmarinterapi förbättrade signifikant det allmänna minnet och det sekundära minnet, bättre än lavendel och kontrollgruppen - För ADHD
Särskilt användbart vid minnesproblem, “hjärndimma” och morgontrötthet - Hur man applicerar
Morgon i diffusorn (5 droppar), inhalation inför prov eller viktiga uppgifter - Lystring! Använd inte på kvällen, det är för aktiverande; var särskilt försiktig vid epilepsi p.g.a. kamferinnehållet!
Lavendel – mot ångest och sömnproblem
Lavendel (Lavendel) är den bästa oljan vid ADHD-relaterad ångest och sömnproblem.
- Vad finns det i?
Linalool (25–45 %)
Linalylacetat (25–50 %) - Vad gör linalool?
Aktiverar GABA-A-receptorer, hjärnans “broms”
Modulerar serotonin, dopamin och noradrenalin
Sänkt kortisol (stresshormon) med 24–38 % - Särskilt användbart vid ADHD
Svårigheter att somna (mycket vanligt vid ADHD)
Ångeststörningar (hos ca 50 % av ADHD-patienterna)
Känslomässig dysreglering och irritabilitet - Viktig meddelande
Lavendel kan det försämra arbetsminnet på kort sikt, därför inte under inlärningsfaser, utan endast på kvällarna eller för avsiktlig avslappning - studera
Silexan (80 mg lavendelolja dagligen som kapsel) verkade jämförbart med lorazepam vid ångeststörningar - Hur man applicerar
Kvällstid i diffusorn (5 droppar), 2 droppar på kudden
Ceder – Syrgasning och fokus
Cederträ (Cedrus atlantica / Juniperus virginianaanvänds traditionellt vid ADHD.
- Vad finns det i?
α-Cedren
β-Cedren
Cedrol - Vad det gör
Förbättrar syresättningen av hjärnan; lugnande; aktiverar parasympatiska nervsystemet - ADHD-specifik
Godfrey (2001) rapporterade positiva effekter hos barn med ADHD;
Cedrol sänkte hjärtfrekvensen och andningsfrekvensen i studier - Hur man applicerar
I diffusorn under lugna aktiviteter; lätt att kombinera med lavendel
Vetiver – jorddoften
Vetiver (Vetiver) har en djup, jordig doft, ofta beskriven som “en lugnare för oroliga själar”.
- Vad det gör
Ångestdämpande; lugnar nervsystemet; förbättrar sömn - ADHD-specifik
Springer et al. (2018) rapporterade förbättringar hos barn med ADHD efter 30 dagars inhalation 3 gånger dagligen - För vem
Särskilt vid hyperaktiv-impulsiv ADHD-variant med inre rastlöshet
Kanel – den enda direkta ADHD-studien
Kanel (Cinnamomum verum) har det unika beviset: den enda kliniska studien direkt på barn med ADHD.
- Vad finns det i?
Zimtaldehyd (60–75 %)
Eugenol (5–10 %) - ADHD-studie (Chen et al., 2008)
Kombination av rehabilitering + kanel-aromaterapi (% kanel, nasal inhalation) under 6 månader visade signifikant bättre resultat i SNAP-IV-frågeformuläret än rehabilitering ensam (p < 0,05) - Mekanism
Zimtaldehydmetaboliter kan ha dopaminerg verkan - Begränsning
Studien har metodologiska svagheter (litet urval, oklar blindning) - Hur man applicerar
Sparsam i diffusorn, kombinerad med andra oljor
Svartpeppar (β-karyofyllen) – mot neuroinflammation
β-karyofyllen av svartpeppar, lavendel och andra växter:
- Aktiverar CB2-cannabinoidreceptorer utan psykoaktiv verkan
- Minskar neuroinflammation (inflammation i hjärnan)
- Intressant med ADHD med samtidiga inflammatoriska tecken eller stämningsproblem
- Lätt att kombinera med andra oljor
Oljorna i översikt – När vilken olja?

Praktisk daglig rutin för ADHD
Morgonrutin (Aktivering + Fokus):
- Diffusor Rosmarin (4 droppar) + pepparmint (2 droppar) i 15–20 minuter
- Eller: Pepparmynta på näsduk, andas in djupt 3 gånger före skolan/jobbet
- Effekt: Vakenhet, minne, koncentration
Lärandefas / Arbetsfas
- Diffusor Pepparmynta (3 droppar) + citron (2 droppar) i 20 minuter, sedan 30 minuters paus
- Lystring! Ingen lavendel under studietiden!
- Effekt: Varaktig uppmärksamhet, fokus
Eftermiddag (Avkoppling efter skolan):
- Diffusor Cederträ (3 droppar) + bergamott (2 droppar) i 20 minuter
- Effekt: Stresshantering, emotionell reglering
Kvällsrutin (Varva ner + Sov):
- Diffusor Lavendel (4 droppar) + Vetiver (2 droppar) 30 minuter före sänggåendet
- Eller: 2 droppar lavendel på kudden
- Effekt: Att somna, sömnkvalitet
Dosering för diffusorn

Viktigt: Vid olfaktorisk adaption märks inte oljan längre efter 20–30 minuter, men den verkar fortfarande. För att receptorerna ska kunna återhämta sig och för att vaneeffekten ska „återställas“ måste pauserna följas!
Vad säger vetenskapen egentligen?

Slutsats
Beviset är mycket lovande men fortfarande begränsad. Mekaniskt sett är det logiskt, men stora kliniska studier på ADHD-patienter saknas fortfarande. Det betyder inte att det inte fungerar, det betyder att vi behöver mer forskning.
Viktiga säkerhetsanvisningar
Särskilda överväganden för ADHD:
Sensorisk överkänslighet
Många personer med ADHD (40–60 %) har ökad luktkänslighet. Börja alltid med halv dos och öka långsamt. Om en doft är obehaglig: sluta omedelbart.
Interaktioner med metylfenidat
- Kanel
Kan dopaminerg effekt ha, vilket teoretiskt möjliggör en förstärkande effekt med MPH, därför behöver hjärtfrekvens och blodtryck övervakas
- Lavendel
Kan ha serotonerg effekt, var försiktig vid samtidig användning av antidepressiva medel
- Rosmarin
Kan verka stimulerande, applicera därför inte strax före sänggåendet
Hos barn
– Under 3 år: Inga eteriska oljor
– 3–6 år: Endast mycket låga koncentrationer (1–2 droppar), väl ventilerat rum
– > 6 år: Normal dosering möjlig – Ingen direkt hudkontakt utan spädning
Allmänna regler:
- Späd alltid med en bärolja (t.ex. oljefri, fraktionerad kokosolja) (2–3 %)
- Använd ett välventilerat utrymme
- Håll en paus (sprid inte ut det hela dagen)
- Vid astma eller andningsproblem: Rådgör med läkare
- Eteriska oljor är ingen ersättning för Metylfenidat eller andra ADHD-behandlingar
FAQ – Vanliga frågor
Kan jag ersätta metylfenidat med eteriska oljor?
Nej. Eteriska oljor är ett komplement, inte en ersättning. Metylfenidat har stark klinisk evidens hos 70–80 % av patienterna. Eteriska oljor kan hjälpa till att lindra kvarvarande symtom, förbättra sömnen eller minska biverkningar (som ångest).
Vilken olja ska jag prova först?
Pepparmynta för koncentration (under dagen) och lavendel för sömn (på kvällen), bäst undersökt, väl tolererad.
Hur lång tid tar det innan jag märker någon effekt?
Vissa effekter (fokus, vakenhet) märks inom några minuter. För varaktiga förbättringar (sömn, humör) bör du använda regelbundet i 2–4 veckor.
Fungerar det även för vuxna med ADHD?
Ja, de flesta studierna har faktiskt genomförts på vuxna. Vuxna med ADHD rapporterar ofta särskilt god effekt på känslomässig dysreglering och sömnproblem.
Måste jag köpa dyra oljor?
Kvalitet är viktigt: Se upp för 100 % 100 % rena eteriska oljor, helst med ett laddningsspecifikt analyscertifikat (GC/MS).
Billiga parfymoljor eller syntetiska doftämnen har ingen terapeutisk effekt och kan, på grund av de syntetiska ingredienserna, vara skadliga för hälsan och orsaka huvudvärk, illamående etc.
Den som vill veta mer om valet och kvaliteten på eteriska oljor hittar information i inlägget „Eteriska oljor – Sökandets odyssé“funnit.
Ett annat inlägg citerar professor Dr. med. habil. Hanns Hatt från Ruhr-Universität Bochum, som i en video säger:„Healing med dofter“förklarar på ett intressant, underhållande och ändå vetenskapligt sätt eteriska oljors verkan på människokroppen.
Ska jag diskutera detta med min läkare?
För aromaterapi (diffuser, inhalation) är det inte absolut nödvändigt. Men om ni vill ta kapslar eller krämer med eteriska oljor, eller om ert barn har ADHD och tar metylfenidat, kontakta er läkare.
Neurobiologiska grunder, molekylära verkningsmekanismer och kliniska bevis för adjuvant terapi med Organik Aromas Nebulizer 3.0
Kombinerad forskningsrapport April 2026
Ämnen Metylfenidat (Ritalin) · Intracerebral farmakodynamik · Adjuvant aromaterapi · Terpener · Receptorer · Signalvägar · Dosrekommendationer
Applikation: Organik Aromas Nebulizer 3.0 (Venturi-kallförstoftning, vattenfri, GC/MS-rena oljor)
ADHD och metylfenidat
Denna rapport undersöker de neurobiologiska grunderna för metylfenidatbehandling vid uppmärksamhetsunderskott-/hyperaktivitetsstörning (ADHD) och utvärderar potentialen hos eteriska oljor som en komplementär behandlingsalternativ. Analysen bygger på 87 vetenskapliga publikationer om intracerebrala metylfenidatkoncentrationer och 159 studier om eteriska oljor i ADHD-kontexten.
Huvudsakliga insikter om metylfenidat
Terapeutiska orala doser (0,25–0,6 mg/kg) ger en striatal dopamintransportörs (DAT) ockupans på >50%, där 20 mg oralt ger 54% och 40 mg 72% DAT-blockad. Maximal hjärnackumulering sker 60–90 minuter efter oral administrering. Plasmanivåer inom det terapeutiska intervallet är 3,5–7,8 ng/ml (vuxna), där 6 ng/ml är associerat med 50% DAT-blockad. Direkta mätningar av metylfenidatkoncentrationer i hjärnvävnad (ng/ml) eller cerebrospinalvätska existerar inte; befintliga data kommer från PET/SPECT-ockupansstudier.
Eteriska oljor som adjuvans
Mekanistisk trovärdighet finns för en adjuvant terapi baserad på kompletterande verkningsmekanismer. Linalool (lavendel) modulerar serotonin-, dopamin- och noradrenalinsystemen via 5-HT1A-receptorer och SERT-bindning. Mentol (pepparmynta) aktiverar TRPM8-kanaler och modulerar GABA-A- samt nikotinerga receptorer, vilket förbättrar uppmärksamhet och vakenhet. β-karyofyllen (svartpeppar) fungerar som en CB2-agonist med antiinflammatoriska egenskaper. 1,8-cineol (rosmarin) förbättrar minnesprestanda och kognitiva funktioner.
Evidensläge
Medan prekliniska och mekanistiska data är lovande, saknas randomiserade kontrollerade studier (RCT) specifikt hos patienter med ADHD. Befintliga mänskliga studier visar kognitiva förbättringar hos friska vuxna (Pepparmynta: förbättrad ihållande uppmärksamhet, n=24; Rosmarin: förbättrat minne, n=144).
Denna rapport ger evidensbaserade doseringsrekommendationer för Organik Aromas Nebulizer 3.0 och omfattande säkerhetsinformation för användning hos patienter med ADHD.
Autismspektrumstörning (ASS)
Autismspektrumstörningen (ASD) är en komplex neurobiologisk utvecklingsstörning med en uppskattad prevalens på 1–2 % globalt. Kärnsymtomen inkluderar nedsättningar i social kommunikation och interaktion samt repetitiva beteendemönster och sensoriska egenheter. Trots intensiv forskning finns inga botande behandlingar för kärnsymtomen; tillgängliga interventioner fokuserar på beteendeterapi och symptomatisk farmakoterapi. Denna rapport undersöker potentialen hos eteriska oljor som en adjuvant behandlingsalternativ vid ASD, baserad på en systematisk analys av 547 vetenskapliga publikationer om ASD-neurobiologi, terpener och klinisk evidens.
ASD:s neurobiologiska heterogenitet manifesteras i störd excitatorisk-inhibitorisk (E/I) balans, dysfunktioner i GABAerga, glutamaterga, serotonerga, dopaminerga och oxytongena system, kronisk neuroinflammation med aktivering av mikroglia, oxidativ stress, förändringar i tarm-hjärna-axeln och synaptiska brister (neuroligin, SHANK-proteiner). Berörda hjärnregioner inkluderar prefrontala cortex, amygdala, lillhjärnan, hippocampus och striatum.
Molekylära analyser visar att specifika terpener från eteriska oljor riktar sig mot ASD-relaterade mål: Linalool (huvudkomponent i lavendel) modulerar GABA-A-receptorer vid bensodiazepinbindningsstället och påverkar serotonerga signalvägar; β-karyofyllen fungerar som en selektiv CB2-receptoragonist med anti-neuroinflammatoriska egenskaper; boswelliasyror (rökelser) aktiverar Nrf2/HO-1-antioxidantvägen och minskar mikrogliaaktivering; 1,8-cineol (eukalyptus) visar ångestdämpande effekter via GABA-A-modulering; α-pinen verkar GABAergt; Cananga odorata (ylang-ylang) påverkar serotonin- och dopaminmetabolismen i prekliniska ASD-modeller.
Den kliniska evidensen vid ASD är begränsad: En randomiserad kontrollerad studie med bergamott visade inga signifikanta anxiolytiska effekter hos ASD-barn. Små pilotstudier rapporterar förbättrad följsamhet under Applied Behavior Analysis (ABA) med lavendel-aromaterapi, ökad delad uppmärksamhet efter aromaterapimassage och möjliga effekter på sömn och ångest. Kvaliteten på evidensen begränsas dock av små urvalsstorlekar, metodologisk heterogenitet och brist på replikering.
För Organik Aromas Nebulizer 3.0 (Venturi-princip, vattenfri kallförångning) utvecklas doseringsrekommendationer anpassade till rumsstorlek: 2–4 droppar för 10–20 m², 4–6 droppar för 20–40 m², 6–10 droppar för 40–60 m², med 15–30 minuters applikationer och 30–60 minuters pauser. ASD-specifika egenheter kräver gradvis introduktion på grund av sensorisk överkänslighet, tidsanpassade protokoll (aktiverande på morgonen, lugnande på kvällen) och strikt kvalitetskontroll (GC/MS-analys).
Säkerhetsaspekter inkluderar försiktighet vid epilepsi (kamferhaltiga oljor), kontraindikation av eukalyptus hos barn under 3 år, potentiella interaktioner med antipsykotika (risperidon, aripiprazol) och särskild hänsyn till sensorisk överkänslighet vid ASD. Mekanistisk rimlighet finns, men högkvalitativa randomiserade kontrollerade studier saknas. Eteriska oljor kan övervägas som en del av en multimodal behandlingsstrategi vid komorbida symtom (ångest, sömnstörningar, sensorisk dysreglering), men ersätter inte evidensbaserade kärninterventioner.
ADHD och metylfenidat
Adjuvant behandling med eteriska oljor vid ADHD – Metylfenidatfarmakodynamik, terpener, receptorer, dosering med Organik Aromas Nebulizer 3.0
Inledning
Attention Deficit Hyperactivity Disorder (ADHD) är en av de vanligaste neuropsykiatriska sjukdomarna hos barn och ungdomar, som drabbar 5–7% av barn och 2,5–4% av vuxna. Metylfenidat (MPH), en hämmare av återupptaget av dopamin och noradrenalin, är den farmakologiska förstahandsbehandlingen och har visat klinisk effekt hos 70–80% av patienterna. [A1], [A2].
Trots metylfenidatets etablerade effekt kvarstår kvarvarande symtom hos en del patienter, biverkningar eller önskemål om kompletterande terapier. Eteriska oljor diskuteras i allt högre grad som ett potentiellt komplementärt behandlingsalternativ, även om den vetenskapliga evidensen hittills är heterogen. [A3], [A4].
Rapportens syfte
Denna rapport har fem huvudmål:
- Kvantifiering av intracerebral metylfenidatkoncentration baserat på PET/SPECT-data, inklusive DAT-ockupans, plasma-hjärnförhållanden och regional fördelning
- Mekanistisk utvärdering Trovärdigheten av en adjuvanserande terapi med eteriska oljor baserat på molekylära verkningsmekanismer
- Evidensbaserad analys Specifika eteriska oljor och deras huvudsakliga aktiva beståndsdelar
- Utveckling av praktiska doseringsrekommendationer för Organik Aromas Nebulizer 3.0
- Utformning av omfattande säkerhetsanvisningar för användning hos ADHD-patienter
Analysen integrerar data från 87 studier om metylfenidat-hjärnkoncentrationer och 159 publikationer om eteriska oljor, där fokus ligger på de 30 mest relevanta publikationerna.
Metylfenidat – Intracerebral Koncentration och Farmakodynamik
DAT-ockupans – Dosis-effekt-samband
Den terapeutiska effekten av metylfenidat är direkt korrelerad med dopamintransportörernas (DAT) ockupans i striatum. PET-studier med [¹¹C]kokain som spårare har etablerat ett tydligt dos-responsförhållande [A9].
Tabell 1: DAT-ockupans efter oral administrering av metylfenidat

Källa: Volkow et al. 1998 [A9]; Mätningar hos friska vuxna
Den uppskattade dosen för 50% DAT-blockad (ED₅₀) är 0,25 mg/kg kroppsvikt [A9]. Terapeutiska viktanpassade doser (0,3–0,6 mg/kg) ger därmed konsekvent ett DAT-mättnadsgrad >50%, vilket betraktas som tröskelvärdet för klinisk effekt [A7], [A9].
Regionalskillnader: DAT-ockupansen varierar mellan striatala subregioner. Spencer et al. (2009) rapporterade följande ockupansvärden för OROS-MPH (40 mg) efter 10 timmar: höger nucleus caudatus 44,3±11,8%, vänster nucleus caudatus 42,7±10,9%, höger putamen 41,6±11,7%, vänster putamen 41,3±13,1%. [A17].
SPECT-studier på ADHD-patienter har visat en signifikant minskning av striatal DAT-bindning under metylfenidatbehandling med [¹²³I]β-CIT. Aster et al. (2021) dokumenterade en 27,6–29,9%minskning av striatalt DAT-bindningspotential (BP) hos barn och ungdomar med ADHD under medicinering. [A25], [A18].
Plasma- kontra hjärnkoncentration
Plasmanivån
Det terapeutiska plasmaområdet för d-treometylfenidat ligger hos vuxna mellan 3,5–7,8 ng/ml [A20], [A24]. En plasmakoncentration på 5,7–6,0 ng/ml är associerad med 50% DAT-blockad [A9], [A13], [A17].
Högre toppkoncentrationer observerades hos pediatriska patienter: 20 ng/ml två timmar efter en dos på 20 mg (0,8 mg/kg). [A20], [A24].
Hjärnaktivitet
PET-studier med [¹¹C]metylfenidat visar att 7,5±1,5% av den intravenöst administrerade aktiviteten når hjärnan [A5], [A14]. Den högsta koncentrationen uppnås i striatum, med ett distributionsvolymförhållande (DVR) mellan basala ganglierna och cerebellum på 2,2–3,3 för d-threo-MPH [A16], [A14].
Kritisk evidenslucka
Direkta mätningar av metylfenidatkoncentrationer i hjärnvävnad (i ng/ml eller nmol/L) eller i cerebrospinalvätska finns inte i den mänskliga litteraturen. All tillgänglig data bygger på PET/SPECT-ockupansmätningar eller tracer-distributionsstudier. [A1], [A2], [A3], [A4], [A5], [A6], [A7], [A8], [A9].
Regional distribution – Striatum and prefrontal cortex
Striatum
Som den primära måletregionen visar striatum den högsta [¹¹C]metylfenidatupptaget. Förbehandling med omärkt metylfenidat (0,5 mg/kg) minskar den striatala spårbindningen med 40 %% [A5]. Clearance från striatum är ~90 minuter, betydligt långsammare än för kokain (20 minuter) [A5].
Prefrontala cortex
Även om DAT-densiteten är lägre i prefrontala cortex (PFC) jämfört med striatum, visar funktionella PET-studier med [¹¹C]racloprid signifikanta dopaminökningar även i prefrontala och temporala kortikala regioner efter metylfenidatadministration [A30], [A22]. Dessa kortikala dopaminökningar korrelerar med långsiktiga symtomförbättringar hos ADHD-patienter [A30].
Noradrenalin-transportör (NET)
Metylfenidat blockerar även noradrenalintransportören med en ED₅₀ på 0,14 mg/kg [A11]. PET-studier med [¹¹C]MRB visar robust NET-ockupans i thalamus och pons 75–195 minuter efter dosering [A11], [A21].
Tidsförlopp och farmakokinetik
Intravenös administrering
Efter intravenös injektion av [¹¹C]metylfenidat når hjärnkoncentrationen sitt maximum inom 8–10 minuter [A3], [A6].
Oral gåva
Den maximala hjärnkoncentrationen efter oral administrering uppnås tydligt fördröjt, med en topp vid 60–90 minuter [A7], [A9], [A3]. Denna fördröjda kinetik förklarar den lägre missbrukspotentialen för orala formuleringar jämfört med intravenösa applikationer [A13].
Retardformuleringar
Depåberedningar (OROS-MPH, DBDS-MPH) visar en ännu långsammare insättande effekt med fördröjd maximal DAT-ockupans, men jämförbara toppkoncentrationer [A13], [A17]. OROS-MPH uppnår signifikant högre DAT-ockupansvärden än DBDS-MPH efter 10 timmar (44,3% vs. 34,8% i höger nucleus caudatus) vid högre plasmakoncentrationer (5,2 vs. 3,8 ng/ml). [A17].
Prekliniska mikrodialysdata
I råttor ökar d-treo-metylfenidat den extracellulära dopamin-koncentrationen i striatum med ungefär% [A16]. Denna massiva dopaminökning demonstrerar den funktionella effekten av DAT-blockad på synaptisk neurotransmission.
Eteriska oljor som stödbehandling – Mekanistiska grunder
Kompletterande verkningsmekanismer för metylfenidat

Bild 1: Molekylära signalvägar för eteriska oljor vid ADHD – Dopaminergt system, noradrenergt system och kompletterande terpeners verkningsmekanismer
Trovärdigheten hos en adjuvant terapi med eteriska oljor bygger på komplementära, icke-redundanta verkningsmekanismer jämfört med metylfenidat. Medan MPH primärt hämmar återupptaget av dopamin och noradrenalin, adresserar eteriska oljekomponenter ytterligare neurobiologiska system.
Mekanistisk komplementaritet:
- GABAerg modulering
Mentol och linalool fungerar som positiva allosteriska modulatorer vid GABA-A-receptorer, vilket förstärker den hämmande neurotransmissionen och kan minska hyperaktivitet. [A19], [A20], [A23]. - Serotonerga effekter
Linalool binder till serotonintransportören (SERT) och modulerar 5-HT1A-receptorer, vilket ger ångestdämpande och humörstabiliserande effekter. [A21], [A23], [A24]. - Antiinflammatoriska signalvägar
Beta-karyofyllen aktiverar CB2-receptorer och minskar neuroinflammatoriska processer via NF-κB- och MAPK-signalvägar, som kan vara förhöjda vid ADHD. [A14]. - TRP-kanalmodulering
Mentol (TRPM8-agonist) och 1,8-cineol (TRPV1/TRPA1-modulator) påverkar neuronal retbarhet och jonflöden oberoende av monoaminerga system [A19].
Receptorer och signalvägar
Tabell 2: Receptormål för eteriska oljekomponenter

LEO = Lavendel eterisk olja; nAChR = nikotinerg acetylkolinreceptor
GABA-A-receptormodulering
Mentol visar koncentrationsberoende bindning till GABA-A-receptorer och förstärker den hämmande neurotransmissionen [A19]. Intressant nog uppvisar lavendelolja ingen affinitet för bensodiazepinbindningsstället på GABA-A-receptorn, vilket tyder på en mer fördelaktig säkerhetsprofil jämfört med bensodiazepiner. [A23].
Serotonin-transportör (SERT)
Linalool och lavendelolja binder till SERT med koncentrationer på 0,08–0,8% in vitro [A23]. Denna SERT-hämning skulle kunna förmedla antidepressiva och ångestdämpande effekter, som är relevanta för ADHD-patienter med komorbida ångestsyndrom (25–40% prevalens).
NMDA-receptonantagonism
Lavendelolja uppvisar dosberoende NMDA-receptorantagonism med ett IC₅₀ på 0,04 µl/ml [A23]. NMDA-receptormodulering är relevant för synaptisk plasticitet och inlärningsprocesser.
Aktivering av CB2-receptorer
β-Caryophyllen är en selektiv CB2-agonist utan psykoaktiva effekter (ingen CB1-aktivering). CB2-aktivering minskar mikrogliainflammation och produktion av proinflammatoriska cytokiner (TNF-α, IL-1β, IL-6), vilket kan förmedla neuroprotektiva effekter. [A14].
Synergistiska potentialer
Kombinationen av metylfenidat med eteriska oljor kan skapa synergistiska effekter genom flera mekanismer:
- Dopamin-GABA-balans
Medan MPH ökar den dopaminerga transmissionen, kan den GABAerga moduleringen av mentol/linalool främja en mer balanserad excitatorisk-inhibitorisk balans och minska hyperaktivitet. - Stressreducering
De ångestdämpande effekterna av linalool via 5-HT1A-receptorer och kortisolreduktion (24–38% minskning i kliniska studier) [A24] skulle kunna förbättra stressreaktiviteten hos ADHD-patienter. - Kognitiv förstärkning
Den kolinerga moduleringen genom mentol (acetylkolinesteras-hämning) [A19] och de minnesfrämjande effekterna av 1,8-cineol kan förstärka de kognitiva förbättringarna av MPH. - Neuroprotektion
De antiinflammatoriska och antioxidativa egenskaperna hos β-karyofyllen och 1,8-cineol kan ge långsiktiga neuroprotektiva effekter.
Viktig begränsning: Dessa synergistiska potentialer är teoretiskt plausibla, men inte validerade genom kliniska studier på ADHD-patienter. Kontrollerade studier av kombinationen av MPH med eteriska oljor saknas helt.
Specifika eteriska oljor – aktiva ingredienser och evidens
Lavendel (Lavandula angustifolia) – Linalool
Huvudsakliga aktiva ingredienser
Linalool (25–45%)
Linalylacetat (25–45%)
1,8-Cineol (1–3%)
Mekanismen
- SERT-bindning
Linalool och lavendelolja binder till serotonin-transportören, vilket kan ge antidepressiva effekter [A23]
- NMDA-antagonism
Lavendelolja uppvisar dosberoende NMDA-receptoblockad (IC₅₀: 0,04 µl/ml). [A23]
- GABA-modulering
Förstärkning av den hämmande neurotransmissionen via GABA-A-receptorer (inte via bensodiazepinbindningsstället) [A20], [A23]
- Monoaminerg effekt
Linalool modulerar dopamin-, noradrenalin- och serotoninsystemen i prekliniska modeller [A21]
Klinisk evidens
- Kognition
Moss et al. (2003) visade hos n=144 friska vuxna att lavendel-aromaterapi försämrade arbetsminnet och saktade ner reaktionstider, men hade en subjektivt lugnande effekt [A27]
- Uppmärksamhet
Colzato et al. (2014) fann att doften av lavendel minskade “attentional blink” (temporala uppmärksamhetsunderskott), vilket tyder på en bredare uppmärksamhetsfördelning [A30]
- Anxiolys
Oral administrering av Silexan (80 mg/dag lavendelolja) reducerade generaliserade ångestsymtom jämförbart med lorazepam [A26]
- Stressreducering
Inhalering av lavendelolja minskade kortisolnivåerna med 24–38% i kliniska studier [A24]
ADHD-specifika data
Inga randomiserade kontrollerade studier hos ADHD-patienter. Fallrapporter beskriver positiva effekter på agitation och sömn hos atypiska barn med ADHD [A8].
Dosering (preklinisk/klinisk)
– Oral: 50–100 µL inneslutna (Kennedy-studien) [A19]
– Inhalation: 4 droppar i 30 ml vatten (doftspridare) [A30]
– Topisk: 1,5% Spädning i basolja [A8]
Säkerhet
Lavendelolja visade ingen förstärkning av cellulär neurotoxicitet in vitro [A23].
Inga allvarliga biverkningar i kliniska studier vid rekommenderade doser.
Rosmarin (Rosmarinus officinalis) – 1,8-Cineol
Huvudsakliga aktiva ingredienser
1,8-Cineol (Eukalyptol, 20–50%)
Kamfer (10–20%)
α-Pinen (10–15%)
Mekanismen
- Kolinerg modulering
Acetylkolinesterashämning, som förstärker kolinerg neurotransmission [A27]
- TRP-kanalaktivering
1,8-Cineol modulerar TRPV1 och TRPA1, vilket påverkar neuronal retbarhet
- Antiinflammatoriskt
Hämning av NF-κB och MAPK-signalvägar, minskning av proinflammatoriska cytokiner
- Antioxidant
Aktivering av Nrf2-signalvägen, ökning av antioxidativa enzymer
Kliniska bevis - Minne
Moss et al. (2003) visade hos n=144 friska vuxna signifikanta förbättringar av det globala minnet och det sekundära minnet efter rosmarinterapi jämfört med kontroll- och lavendelgrupperna [A27]
- Kognitiv prestation Exponering för rosmarindoft korrelerade med förbättrad kognitiv prestation [A14]
- Vakenhet: Rosmarinen ökade vakenhet och aktivitet i hjärnan [A29]
ADHD-specifika data
Rosmarin nämns i översiktsartiklar som potentiellt hjälpsamt vid ADHD [A10], [A4], [A5], men kontrollerade kliniska studier saknas hos ADHD-patienter.
dosering
– Inhalation: Omgivningsexponering i testmiljö (koncentration ej specificerad) [A27]
– Inga standardiserade doseringsrekommendationer för ADHD tillgängliga
Säkerhet
Rosmarin anses vara säkert vid inhalativ användning i vanliga koncentrationer.
Varning för patienter med epilepsi på grund av kamferinnehållet.
Pepparmynta (Mentha × piperita) – Mentol
Huvudsakliga aktiva ingredienser
Mentol (30–50%)
Menton (10–30%)
1,8-Cineol (5–10%)
Mekanismen
- TRPM8-aktivering
Mentol är en potent TRPM8-agonist (EC₅₀ ~30 µM), vilket medierar köldkänsla och neuronal modulation [A19]
- GABA-A-modulering
Koncentrationsberoende bindning och positiv allosterisk modulering [A19]
- Nikotinreceptorer
Modulering av nikotinerga acetylkolinreceptorer, påverkan av kolinerg transmission [A19]
- Acetylkolinesterashämning
Pepparmyntsolja hämmar signifikant acetylkolinesteras, vilket förstärker kolinerg neurotransmission [A19]
Kliniska bevis - Varaktig uppmärksamhet
Kennedy et al. (2018) visade i en dubbelblind, crossover-studie (n=24 friska vuxna) att 100 µL pepparmyntsolja förbättrade prestationen i testet Rapid Visual Information Processing (RVIP) 1–3 timmar efter intag [A19]
- Trötthetsreducering Båda doserna (50 µL och 100 µL) minskade trötthet och förbättrade seriel subtraktion efter 3 timmar [A19]
- Uppmärksamhetskontroll: Colzato et al. (2014) fann att pepparmintsarom förstärkte “attentional blink”, vilket tyder på mer fokuserad (mindre spridd) uppmärksamhet [A30]
ADHD-specifika data
Inga RCT:er hos ADHD-patienter. Pepparmynta nämns som potentiellt hjälpsam i översiktsartiklar om ADHD. [A4], [A5], [A29].
dosering
– Oralt (inkapslat): 50–100 µL ren olja [A19]
– Inhalation: 4 droppar i 30 ml vatten (doftspridare) [A30]
Säkerhet
Pepparmyntolja är säker vid inhalatorisk och oral användning i rekommenderade doser.
Försiktighet hos spädbarn och små barn (risk för laryngospasm vid direkt nasal administrering).
Svartpeppar (Piper nigrum) – β-karyofyllen
Huvudsakliga aktiva ingredienser
β-Caryophyllen (–35)
Limonen (15–25%)
Sabinen (10–20%)
Mekanismen
- CB2-agonism
β-Caryophyllen är en selektiv CB2-receptoragonist (Ki: 155±4 nM) utan CB1-aktivering (inga psykoaktiva effekter). [A14]
- Antiinflammatoriskt
Reduktion av TNF-α, IL-1β, IL-6 via NF-κB-hämning [A14]
- Mikrogliamodulering
Omprogrammering från M1 till M2 mikrogliatyper, minskning av neuroinflammatoriska processer
- Neuroprotektiv
Minskning av oxidativ stress genom Nrf2-aktivering
Klinisk evidens
- Preklinisk
β-Caryophyllen visade ångestdämpande effekter i djurmodeller, möjligen genom modulering av neuronal aktivitet i den centrala amygdalan. [A14]
- Antiinflammatoriskt
In vitro- och in vivo-studier visar robusta antiinflammatoriska effekter via CB2-aktivering
ADHD-specifika data
Inga kliniska studier hos ADHD-patienter. Mekanistisk plausibilitet baserad på neuroinflammatoriska hypoteser vid ADHD.
dosering
Inga standardiserade doseringsrekommendationer för inhalation vid ADHD finns tillgängliga.
Säkerhet
β-Caryophyllen anses vara säkert (GRAS-status enligt FDA).
Inga kända allvarliga biverkningar vid vanliga doser.
Vetiver (Chrysopogon zizanioides)
Huvudsakliga aktiva ingredienser
Vetiverol
Khusimol
α-Vetivon
β-Vetivon
Mekanismen
- Ångestdämpande
Präkliniska studier visar ångestdämpande effekter, möjligen via modulering av amygdala-aktivitet [A14]
- Lugnande
Traditionellt använd för att lugna nervsystemet
Klinisk evidens
- ADHD-specifik
Springer et al. (2018) rapporterar att vetiverinhalation (3 gånger dagligen i 30 dagar) visade förbättringar av hjärnfunktion och beteende hos barn. [A29]
- Begränsningar
Inga kontrollerade randomiserade studier; bevisen baseras på fallrapporter och traditionell användning
ADHD-specifika data
Godfrey (2001) nämner Vetiver som potentiellt hjälpsamt vid ADHD [A3], [A4], [A5], men detaljerade kliniska data saknas.
dosering
– Inhalation: 3 gånger dagligen i 30 dagar (specifik koncentration ej angiven) [A29]
Säkerhet
Vetiver anses vara säkert vid inandning. Inga kända allvarliga biverkningar.
Zeder (Cedrus spp.)
Huvudsakliga aktiva ingredienser
α-Cedren
β-Cedren
Thujopsen
Cedrol
Mekanismen
- Syresättning
Traditionellt använd för att förbättra syresättningen i hjärnan [A29]
- Lugnande
Lugnande egenskaper i traditionell användning
Klinisk evidens
- ADHD-specifik
Godfrey (2001) nämner cederträ som potentiellt hjälpsamt vid ADHD [A3], [A4], [A5]
- Begränsningar
Inga kontrollerade kliniska prövningar; bevisen baseras på traditionell användning och anekdoter
ADHD-specifika data
Springer et al. (2018) nämner cederträ för hjärnans syresättning vid ADHD [A29], men kvantitativa data saknas.
dosering
Inga standardiserade doseringsrekommendationer tillgängliga.
Säkerhet
Cederoljebruk anses vara säkert vid inhalativ användning vid normala koncentrationer.
Kanel (Cinnamomum verum) – Kanelaldehyd
Huvudsakliga aktiva ingredienser
Zimtaldehyd (60–75%)
Eugenol (5–10%)
Mekanismen
- Dopaminerg modulering
Allylbenzen och propenylbenzen i kanel är förstadier till amfetamin och omvandlas efter metabolism till amfetamin, vilket har en mentalt stimulerande effekt [A2]
- TRPA1/TRPV1-aktivering
Zimtaldehyd aktiverar TRPA1- och TRPV1-kanaler
- Antimikrobiell
Starka antimikrobiella egenskaper
Kliniska bevis - ADHD-specifik
Chen et al. (2008) genomförde en studie med kanel-aromaterapi hos barn med ADHD. Kombinationen av rehabilitering och kanel-aromaterapi (1% kanel, 1 g i 100 ml vatten, nasal inhalation) visade signifikant bättre resultat i SNAP-IV-enkäten (58±2,6 vs. kontroll) och i aktivitets-skalor (102±5,8) efter 6 månader jämfört med enbart rehabilitering (p<0,05). [A2], [A1]
ADHD-specifika data
Chen-studien är en av få kliniska studier med eteriska oljor specifikt på barn med ADHD, men med metodologiska begränsningar (inga uppgifter om randomisering, blindning, urvalsstorlek).
dosering
– Inhalation: 1% Kanel (1 g i 100 ml vatten) i ett 50 m² terapirum med elektrisk ljusstyrning [A2]
Säkerhet
Kanelolja kan orsaka hud- och slemhinneirritationer vid höga koncentrationer.
Försiktighet kan krävas för känsliga personer.
Den dopaminerga stimuleringen genom kanelmetaboliter kräver ytterligare undersökningar gällande interaktioner med metylfenidat.
Ytterligare förtecknade eteriska oljor vid ADHD
Baserat på en aktuell litteraturgenomgång har följande ytterligare eteriska oljor identifierats som kan vara relevanta för ADHD genom neurobiologiskt plausibla mekanismer.
Melissa officinalis (Citronmeliss) – Rosmarinsyra, flavonoider
Aktiva ingredienser
Rosmarinsyra
Flavonoider (Luteolin, Apigenin)
Flyktiga oljor (Citral, Linalool, Geraniol)
Receptorer och verkningsmekanismer
- Kolinerga receptorer: Nikotinerga (nAChR) och muskarinerga (mAChR) bindning
– GABAerg modulering: Rosmarinsyra hämmar GABA-transaminas → ökad GABA-tillgänglighet
– Acetylkolinesterashämmning: Förbättrad kolinerg neurotransmission i prefrontala cortex
Klinisk evidens
- RCT (n=20, dubbelblind, korsade mönsterstudier)
Enstaka doser om 300/600/900 mg peroralt visade vid 600 mg signifikant förbättring av uppmärksamhetsnoggrannhet (Accuracy of Attention) samt minnes- och vakenhetseffekter. [A31]
- Kombinationsstudie (barn, 7 veckor)
Valeriana-melissaextrakt-kombination förbättrade signifikant hyperaktivitet, koncentrationssvårigheter och impulsivitet hos grundskolebarn [A32]
- Antistresscrossover
Standardiserade citronmeliss-preparat i livsmedelsform minskade kognitiv överbelastning och förbättrade humöret hos friska vuxna [A33]
Dosering (Organik Aromas Nebulizer 3.0)
– 3–5 droppar citronmelissolja (GC/MS-ren) för ett rum på 15–25 m²
– Användning: 30 min före koncentrationsperioder, 2–3 gånger dagligen
– Kombination med lavendel (2:1) rekommenderas för kvällen/avkoppling
Säkerhet
Lätt att smälta
Möjlig växelverkan med sköldkörtelmedicin (TSH-suppression vid hög oral dosering)
Inandning anses vara säker.
Eukalyptus (Eucalyptus globulus / radiata) – 1,8-Cineol
Aktiva ingredienser
1,8-Cineol (Eukalyptol, 70–85%)
α-Pinen
Limefrukter
Receptorer och verkningsmekanismer
– GABA-A/Bensodiazepin-receptor: Modulerande aktivitet (ångestlindrande, sederande vid hög dos)
– Kolinerga banor: 1,8-Cineol hämmar acetylkolinesteras → förbättrad uppmärksamhet och arbetsminne
– Antioxidativ/Neuroprotektiv: Minskning av oxidativ stress i neuronala celler
Klinisk evidens
– Neuroprotektive Reviews listar Eucalyptus globulus som en växt med kognitiva effekter i djur- och humanstudier [A34]
– 1,8-Cineol är också den huvudsakliga aktiva substansen i rosmarin (redan inkluderad i rapporten), eukalyptus ger högre ren koncentration
– Inhalationsstudier visar förbättrad mental klarhet och koncentration (jämförande studier med rosmarin)
Dosering (Organik Aromas Nebulizer 3.0)
– 2–3 droppar för rum 15–25 m² (intensiv doft, dosera sparsamt)
– Användning: Morgon/Middag för koncentrationsfaser, max 20 min
- Kontraindikation
Inte för barn under 3 år
Används inte i kombination med astmamediciner utan läkares rådgivning
Säkerhet
Stark olja, applicera inte outspädd på huden
Barn < 3 år: kontraindicerat
Epileptiker: Försiktighet
Ingefära (Zingiber officinale) – Ingefäraolen, Zingiberen
Aktiva ingredienser
6-Gingerol
6-Shogaol
Zingiberen
β-Sesquiphellandren
Receptorer och verkningsmekanismer
Serotonin-5-HT3-receptorantagonism: Anxiolytisk komponent
– Dopaminerg: Gingerol modulerar dopaminerg aktivitet i striatum (djur)
– Antioxidativ/Anti-neuroinflammatorisk: NF-κB-hämning, minskning av TNF-α
Klinisk evidens
– Preklinisk: Ingefäraextrakt förbättrade kognitiv funktion i ADHS-djurmodeller
– Neuroprotektiva egenskaper genom antioxidativa mekanismer väl dokumenterade
– Direkt jämförande ADHS-RCT saknas fortfarande i litteraturen
Dosering (Organik Aromas Nebulizer 3.0)
– 2–3 droppar för rum 15–25 m²
– Kombination med rosmarin (1:1) för kognitiv aktivering
Dosrekommendationer för Organik Aromas Nebulizer 3.0
Tekniska grunder: Venturiprincipen och förstoftning
De Organik Aromas Nebulizer 3.0* använder Venturi-principen för vattenfri kallförstoftning av eteriska oljor. Denna metod erbjuder flera fördelar:
Tekniska egenskaper
- Kallförstoftning
Ingen värmepåverkan, bevarande av alla flyktiga komponenter och värmekänsliga aktiva ingredienser
- Vattenfri
Direkt nebulisering av rena eteriska oljor utan utspädning, högre koncentration av aktiva ämnen i inomhusluften
- Partikelstorlek
Framställning av mikropartiklar (1–5 µm) som kan nå djupt in i luftvägarna och passera blod-hjärnbarriären
- GC/MS-certifierade rena oljor
Användning av analytiskt testade oljor utan tillsatser, basoljor eller syntetiska doftämnen
ca 110 till 120 USD (med nät-/batteridrift
- kostnadsfri leverans över hela världen
- Tänk på tull- och importskatter!
Farmakokinetiska överväganden
Den inhalativa upptagningen av eteriska oljor sker via två huvudsakliga vägar:
1. Olfaktorisk väg
Direkt koppling från luktepitelet till det limbiska systemet (amygdala, hippocampus) och prefrontala cortex via luktbulben
2. Pulmonal väg
Absorption över alveolarmembranen till systemkretsloppet, passage av blod-hjärnbarriären [A29]
Rumstorlek och droppmängd
Grundformel
Droppmängden bör anpassas till rummets storlek för att uppnå en terapeutiskt effektiv men inte överdriven koncentration.
Tabell 3: Dosrekommendationer efter rumsstorlek

*Antagande: Rumshöjd 2,5 m **Uppskattad koncentration baserad på ~1 mg eterisk olja per droppe och fullständig förångning
Beräkningsunderlag
– 1 droppe eterisk olja ≈ 0,05 ml ≈ 40–50 mg (beroende på densitet)
– Målsättning: 80–200 µg/m³ (baserat på studier med omgivande aromaterapi)
– Formel: Droppar = (Rymdvolym × Målkoncentration) / (Massa per droppe × 1000)
Användningstid och pauser
Sessionstid
- Akut användning
15–30 minuter per session
- Kontinuerlig bakgrundsdiffussion
30–60 minuter med pauser
Pausintervall
Den olfaktoriska adaptationen (vanan) inträder efter 15–30 minuters kontinuerlig exponering.
För att bibehålla terapeutiska effekter rekommenderas pauser:
Tabell 4: Användnings- och pauschema

Olfaktorisk adaptation
Efter 20–30 minuters kontinuerlig exponering minskar den medvetna uppfattningen av doften, men neurobiologiska effekter kan kvarstå.
Pausar på 30–60 minuter möjliggör en återkänslighet av luktreceptorerna.
Dagtider och användningsscenarier
Valet av eteriska oljor bör anpassas efter tid på dygnet och specifika ADHD-symtom:
Tabell 5: Oljeval baserat på tid på dygnet

Användningsfall:
- Schulvorbereitung/Morgenroutine
– Öl: Rosmarin eller pepparmynta
– Mål: Aktivering, vakenhet, fokus. Dosering: 6–8 droppar i 20 minuter.
– Tidpunkt: 30–60 minuter före skolstart - Läxor / Lärandefaser
– Öl: Pepparmynta (varaktig uppmärksamhet) eller rosmarin (minne)
– Mål: Koncentration, arbetsminneDosering: 6–8 droppar i 30 minuter, sedan 30 minuters paus
– Tidpunkt: Under läxor eller studiepass - Hantera hyperaktivitet:
– Öl: Lavendel eller Vetiver
– Mål: Lugnande, minskning av motorisk rastlöshet Dosering: 5–7 droppar i 30 minuter
– Tidpunkt: Vid akut hyperaktivitet eller agitation - Sömnhygien
– Öl: Lavendel
– Mål: Insomningshjälp, minskning av insomningsproblem (vanligt vid ADHD)Dosering: 4–6 droppar i 15 minuter
– Tidpunkt: 30–60 minuter före sänggåendet
Öl-specifika doseringar
Tabell 6: Oljespecifika doseringsrekommendationer

Blandningar Kombinationen av flera oljor kan ge synergistiska effekter:
- Fokus-Blend (Förmiddag)
3 droppar rosmarin + 3 droppar pepparmynta + 2 droppar kanel - Lugnande blandning (kväll)
4 droppar lavendel + 3 droppar vetiver + 2 droppar ceder - Balans-Blend (eftermiddag)
3 droppar pepparmynta + 3 droppar lavendel + 2 droppar svartpeppar
Viktigt
Håll det totala antalet droppar inom det rekommenderade intervallet för blandningar (6–8 droppar för 20 m²).
Säkerhetsanvisningar och kontraindikationer
Allmän säkerhet vid inandning
Grundprinciper
renhet
Använd endast GC/MS-certifierade, 100% rena eteriska oljor utan tillsatser
Spädning
Vid inandning via en diffusor krävs ingen ytterligare spädning; vid topisk användning späd alltid med en basolja (1–3%)
ventilation
Rummet bör inte vara helt tät; säkerställ regelbunden tillförsel av frisk luft
Individuell tolerans
Börja med låga doser och öka gradvis
Vanliga biverkningar (milda)
– Huvudvärk vid för hög koncentration eller för lång exponering
– Slemhinneirritation (näsa, svalg) hos känsliga personer
– Illamående av mycket intensiva dofter (t.ex. kanel, svartpeppar)
– Allergiska reaktioner (sällsynta, <1% av befolkningen)
Åtgärder vid biverkningar
– Stäng av diffusorn omedelbart och vädra ur rummet
– Vid huvudvärk: Frisk luft, vätska
– Vid slemhinneirritation: Avbryt inhalation, vid behov inhalera koksaltlösning
– Vid allergiska reaktioner: Avsluta exponering, eventuellt antihistaminikum, vid svåra reaktioner läkarvård
ADHD-specifika överväganden
Särskilda sårbarheter hos ADHD-patienter:
Sensorisk överkänslighet
40–60% der ADHD-Patienten zeigen sensorische Verarbeitungsstörungen. Intensive Düfte können als unangenehm oder überwältigend empfunden werden.
- Rekommendation
Börja med mycket låga doser (50% av standarddosen), öka gradvis
Komorbida ångeststörningar
25–40% av ADHD-patienter har komorbida ångestsyndrom.
Aktiviserande oljor (rosmarin, pepparmynta) kan förstärka ångest.
- Rekommendation
Vid komorbida åkommor med ångest, använd primärt ångestdämpande oljor (lavendel, vetiver).
Sömnstörningar
50–70% av ADHD-patienter har sömnproblem. Aktiverande oljor på kvällen kan försämra sömnen ytterligare.
- Rekommendation
Strikt separation: aktiverande oljor endast på förmiddagen/eftermiddagen, lugnande oljor på kvällen
Impulsivitet
ADHD-patienter kan vara benägna att använda för höga doser.
- Rekommendation
Tydliga doseringsanvisningar, eventuellt övervakning av föräldrar/vårdnadshavare
Interaktioner med metylfenidat
Teoretiska interaktioner:
Dopaminerg förstärkning (kanel)
– Kanelmetaboliter kan omvandlas till amfetamin [A2]
Teoretisk risk för additiva dopaminerg effekter med metylfenidat
- Rekommendation
Använd endast kanel i låga doser, noggrann övervakning av överstimulering (takykardi, agitation, sömnlöshet)
Serotonerg modulering (lavendel)
Linalool binder till SERT [A23]
Teoretisk risk för serotonergt syndrom vid samtidig användning av SSRI (ofta vid komorbid depression/ångest).
- Rekommendation
Vid samtidig behandling med SSRI bör lavendel endast användas i måttliga doser, med övervakning av symtom på serotonergt syndrom (agitation, förvirring, takykardi, hypertermi).
Sedering (Lavendel, Vetiver)
– Sederande oljor kan minska vakenheten
Kontraproduktivt för metylfenidatets stimulerande effekt
- Rekommendation
Använd lugnande oljor primärt på kvällen, inte under skoltid eller vid aktiviteter som kräver uppmärksamhet
- Inga kända farmakokinetiska interaktioner
Eteriska oljor metaboliseras primärt via CYP2D6, CYP3A4
Metylfenidat metaboliseras via karboxylesteras 1 (CES1)
Ingen överlappning av metaboliseringsvägar, därför inga förväntade farmakokinetiska interaktioner
Viktigt
Även om det saknas dokumenterade interaktioner bör kombinationen av metylfenidat med eteriska oljor ske under läkarövervakning, särskilt vid behandlingsstart.
Kontraindikationer och försiktighetsåtgärder
Absolut kontraindikation
– Känd allergi mot specifika eteriska oljor eller deras beståndsdelar
– Svår astma (risk för bronkospasm)
– Spädbarn <6 månader (risk för laryngospasm, särskilt med mentol)
Relativ kontraindikation (försiktighet krävs)
– Epilepsi: Försiktighet med rosmarin och kanel (kamfer, kanelaldehyd kan sänka kramptröskeln)
– Graviditet/amning: Många eteriska oljor har inte studerats tillräckligt under graviditet/amning
– Astma (mild-måttlig): Låga doser, tät uppföljning av luftvägssymtom
– Små barn (6 månader – 3 år): Reducerade doser (25–50% av vuxendosen)
Doseringsjusteringar beroende på ålder:

Kvalitetskriterier för eteriska oljor
- GC/MS-certifikat:
Varje sats måste analyseras med gaskromatografi/masspektrometri - Botaniska namnet
Vetenskapligt namn måste anges (t.ex. Lavandula angustifolia, inte bara “lavendel”) - Kemotyp
Vid oljor med olika kemotyper (t.ex. rosmarin) måste detta specificeras - Ursprung
Ursprungsland och extraktionsmetod (destillation, kallpressning) bör dokumenteras - Inga tillsatser
100% ren, utan syntetiska doftämnen, basoljor eller konserveringsmedel
Lagring och hållbarhet
– Förvaras mörkt och svalt (15–20°C)
– Skydda mot ljus (bruna eller blå glasflaskor)
– Förbrukas inom 6 till 12 månader efter öppning (oxidation)
– Citrixoljor har kortare hållbarhet (6 månader), hartser längre (2 till 3 år)
Diskussion och kliniska implikationer
Evidensluckor och forskningsbehov
Kritisk bedömning av evidensläget:
Denna analys visar en betydande diskrepans mellan mekanistisk trovärdighet och kliniska bevis för eteriska oljor som komplementär ADHD-behandling.
Styrka på tillgänglig evidens
- Robusta mekanistiska data
Receptorbindningsstudier, signalvägsanalyser och prekliniska modeller visar tydliga neurobiologiska effekter av eteriska oljekomponenter [A19], [A20], [A21], [A23] - Kognitiva effekter hos friska
Kontrollerade studier på friska vuxna visar mätbara effekter på uppmärksamhet (pepparmynta, n=24) [A19] och minne (rosmarin, n=144) [A27] - Komplementära verkningsmekanismer
Eteriska oljors molekylära mål (GABA-A, CB2, TRP-kanaler, SERT) skiljer sig från metylfenidats (DAT/NET), vilket antyder synergistiska potentialer
Svagheter och evidensluckor
- Brist på ADHD-specifika RCT:er Med undantag av Chen-studien om kanel [A2] finns inga randomiserade, placebokontrollerade studier hos ADHD-patienter
- Metodologiska begränsningar: Befintliga studier har små stickprov (n=24–144), korta observationsperioder (enskilda sessioner) och saknar ofta blindning
- Inga långtidsdata: Effekter och säkerhet vid kronisk användning (> 6 månader) har inte studerats
- Studier saknas för dosbestämning: Optimala doser, koncentrationer och användningsscheman för ADHD är inte etablerade
- Inga kombinationsstudier: Interaktioner och synergistiska effekter med metylfenidat har inte studerats systematiskt
Specifika forskningsluckor:

Praktisk genomförande
Riktlinjer för klinisk praxis:
Trots bristen på evidens kan en adjuvant terapi med eteriska oljor övervägas hos ADHD-patienter under följande förutsättningar:
1. Tydlig indikation
- Restsymtom vid optimerad metylfenidatterapi
- Patientens, familjens önskemål om kompletterande metoder
- Specifika symptom som kan behandlas med eteriska oljor
(Sömnstörningar → Lavendel; Minnesproblem → Rosmarin)
2. Strukturerat tillvägagångssätt
- Grundbedömning
Dokumentation av ADHD-symtomatik med validerade skalor (SNAP-IV, Conners, ADHS-SB) - Introduktionsfas
Börja med en enda olja i låg dos (50% av standarddosen) i 2 veckor - Övervakning
Veckovis symtomregistrering, biverkningsövervakning - Titrering
Stegvis dosökning vid god tolerans - Utvärdering
Efter 4 till 6 veckor utvärdering av effekt; vid utebliven nytta avbrytande
3. Dokumentation
- Att föra en “aromaterapi-dagbok” med olja, dosering, tidpunkt, symtom, biverkningar
- Regelbundna samråd med behandlande läkare
4. Realistiska förväntningar
- Eteriska oljor är ingen ersättning för evidensbaserade ADHD-behandlingar (metylfenidat, beteendeterapi)
- Förväntad effektstorlek: liten till måttlig (Cohens d: 0,2–0,5)
- Primär nytta: Förbättring av specifika symtom (sömn, ångest, stämningsläge), inte kärnsymtom
Begränsningar
Analysens begränsning:
- Indirekta bevis
De flesta slutsatser baseras på extrapolation av studier på friska vuxna till ADHD-patienter - Publikationsbias
Positiva studier publiceras oft; negativa studier om eteriska oljor kan vara underrepresenterade - Heterogenitet
Olika oljor, doser, administrationsvägar och utfallsmått försvårar metaanalyser - Brist på plasma-/hjärnspegling
Det finns inga data för plasmakoncentrationer i hjärnan efter inandning av eteriska oljekomponenter hos människor. - Mekanisk osäkerhet
De exakta mekanismerna genom vilka inhalerade eteriska oljor medierar CNS-effekter (olfaktoriska kontra systemiska) är inte helt klarlagda
Begränsningen av doseringsrekommendationerna
– Baserat på extrapolationer från ett fåtal studier och traditionell användning
– Individuell variabilitet (rumsluftcirkulation, andningsfrekvens, metabolism) ej beaktad
– Inga farmakokinetiska data om dos-koncentrationssambandet
Slutsats
Denna omfattande analys integrerar data från 87 studier om intracerebrala metylfenidatkoncentrationer och 159 publikationer om eteriska oljor i ADHD-kontexten. Huvudresultaten kan sammanfattas enligt följande:
Metylfenidat hjärnkoncentration
Terapeutiska orala doser av metylfenidat (0,25–0,6 mg/kg) ger en DAT-ockupans på >50%i striatum, med ett välkarakteriserat dos-responsförhållande (20 mg → 54%, 40 mg → 72% DAT-blockad) [A9].
Maximal hjärnkoncentration uppnås 60–90 minuter efter oral administrering [A7], [A9].
Plasmanivåer inom det terapeutiska intervallet ligger på 3,5 – 7,8 ng/ml (vuxna), där 6 ng/ml är associerat med 50% DAT-blockad [A9], [A17].
Kritisk evidenslucka
Direkta mätningar av metylfenidatkoncentrationer i hjärnvävnad (ng/ml) eller cerebrospinalvätska finns inte; all data baseras på PET/SPECT-ockupationsmätningar.
Adjuvant terapi med eteriska oljor
En kompletterande behandling med eteriska oljor är mekanistiskt plausibel baserat på kompletterande verkningsmekanismer.
– Linalool modulerar serotonin-, dopamin- och noradrenalinsystemen [A21], [A23];
– Mentol aktiverar TRPM8-kanaler och GABA-A-receptorer [A19];
– β-Caryophyllen verkar som en CB2-agonist med antiinflammatoriska egenskaper [A14];
– 1,8-Cineol förbättrar kognitiva funktioner [A27].
Dessa mekanismer är komplementära till DAT/NET-blockeringen av metylfenidat och kan ge synergistiska effekter.
Däremot: Den kliniska evidensen hos ADHD-patienter är svag; kontrollerade RCT saknas till stor del.
Specifika oljor och aktiva ingredienser
- Pepparmynta (Mentol)
Starkaste bevis för förbättrad uthållighetsuppmärksamhet (n=24 RCT) [A19] - Rosmarin (1,8-Cineol)
Minnesförbättring (n=144) [A27] - Lavendel (Linalool)
Anxiolys, förbättrad sömn, men nedsatt arbetsminne [A27], [A23] - Svartpeppar (β-karyofyllen)
Antiinflammatorisk, preklinisk ångestdämpande effekt [A14] - Vetiver, cederträ
Svaga bevis, primärt traditionell användning [A3], [A4], [A29] - Kanel (kanelaldehyd)
Enda ADHD-specifika kliniska studien, men metodologiska begränsningar [A2]
Doserings rekommendationer för Organik Aromas Nebulizer 3.0
Evidensbaserade doseringsrekommendationer har utvecklats, anpassade till rumsstorlek (3–18 droppar för 10–60 m²), tid på dygnet (aktiverande oljor på morgonen, lugnande på kvällen) och specifika symtom.
Användningstid: 15–30 minuter per session med 30–60 minuters pauser för att undvika luktadaption.
Viktigt: Dessa rekommendationer baseras på extrapolering från ett litet antal studier och kräver individuell anpassning.
Säkerhetsinstruktioner
Eteriska oljor är generellt säkra vid inhalationsanvändning i rekommenderade doser.
Specifika försiktighetsåtgärder för patienter med ADHD inkluderar:
- Hänsyn till sensorisk överkänslighet (40–60% av ADHD-patienterna)
- Undvik stimulerande oljor vid komorbid ångest
- strikt åtskillnad av aktiverande (förmiddag) och sedativa (kväll) oljor
- Teoretiska interaktioner med metylfenidat (dopaminerg förstärkning genom kanel, serotonerg modulering genom lavendel) kräver övervakning
- Absolut kontraindikation
– känd allergi
– svår astma
– Spädbarn <6 månader
Slutlig bedömning
Eteriska oljor utgör en lovande men otillräckligt validerade adjunkterande behandlingsalternativ vid ADHD. Den mekanistiska plausibiliteten är stark, men den kliniska evidensen är svag.
En applikation kan övervägas för informerade patienter/familjer under medicinsk övervakning, men bör aldrig evidensbaserade terapier ersätter.
Brådskande behov av forskning finns för randomiserade, placebokontrollerade studier hos ADHD-patienter, långtids datasäkerhet och kombinationsstudier med metylfenidat.
Autismspektrumstörning (ASS)
Neurobiologiska grunder för ASD, terpener som CB2/GABA-A/Nrf2-modulatorer, klinisk evidens och dosering med Organik Aromas Nebulizer 3.0
Inledning
Autismspektrumstörningen (ASS) är en av de vanligaste neurobiologiska utvecklingsstörningarna, med en uppskattad global prevalens på 1 till 2 % och en ökande diagnostik under de senaste två decennierna [B1], [B2].
ASD kännetecknas av en uttalad klinisk och neurobiologisk heterogenitet, som manifesteras i ett brett spektrum av symtomuttryck, från högfungerande individer med isolerade brister i social kommunikation till personer med svåra intellektuella funktionsnedsättningar och komorbida neurologiska sjukdomar. [B3], [B4].
Kärnsymptomatiken enligt DSM-5 omfattar två huvuddomäner: ihållande brister i social kommunikation och social interaktion, samt begränsade, repetitiva beteendemönster, intressen eller aktiviteter [B5]. Zusätzlich zeigen bis zu 90 % der Betroffenen sensorische Verarbeitungsstörungen, die sich als Hyper- oder Hyposensitivität gegenüber sensorischen Stimuli äußern [B6], [B7].
Komorbida symtom som ångeststörningar (40–50 %), sömnstörningar (50 till 80 %), uppmärksamhets-/hyperaktivitetsstörning (30 till 50 %) och gastrointestinala besvär (30 till 70 %) påverkar livskvaliteten avsevärt. [B8], [B9], [B10].
Neurobiologisk heterogenitet och patofysiologisk konvergens
Den etiologiska heterogeniteten av ASD omfattar genetiska faktorer (ärftlighet 70 till 90 %), de novo-mutationer i synaptiska gener (SHANK3, NLGN3/4, NRXN1), kopienummervariationer, epigenetiska modifieringar och prenatala miljöfaktorer [B11], [B12], [B13].
Trots denna mångfald konvergerar de patofysiologiska mekanismerna mot gemensamma neurobiologiska slutpunkter: störd exitatorisk-inhibitorisk (E/I) balans, synaptisk dysfunktion, neuroinflammation, oxidativ stress och förändringar i tarm-hjärna-axeln B14, [B15], [B16].
Hypotesen om E/I-obalans postulerar att en obalans mellan excitatorisk (glutamaterg) och inhiberande (GABAerg) neurotransmission förklarar centrala ASD-symtom [B17], [B18].
Bildstudier och postmortemananalyser visar regionala förändringar i GABA- och glutamatkoncentrationer, förändrad expression av GABA-A- och NMDA-receptorer samt dysfunktion i GABAerga interneuron [B19], [B20], [B21].
Denna obalans påverkar särskilt prefrontala cortex (exekutiva funktioner, social kognition), amygdala (känslomässig bearbetning, social perception), lillhjärnan (sensomotorisk integration, kognitiva processer) och hippocampus (minne, kontextuell bearbetning) [B22], [B23], [B24].
Neuroinflammation utgör en annan konvergent mekanism: postmortala studier visar mikroglia- och astrocytaaktivering i ASD-hjärnor, prekliniska modeller (t.ex. maternal immunaktivering, valproinsyreinducerad ASD) demonstrerar ökade proinflammatoriska cytokiner (TNF-α, IL-6, IL-1β) och störd synaptisk utveckling genom kronisk neuroinflammation [B25], [B26], [B27].
Oxidativ stress med ökat reaktivt syre (ROS), minskat glutation (GSH) och mitokondriell dysfunktion har påvisats i flera ASD-kohorter. [B28], [B29].
Tarm-hjärna-axeln spelar en alltmer erkänd roll: patienter med ASD uppvisar ofta dysbios med förändrade firmicutes/bacteroidetes-förhållanden, minskad mikrobiell mångfald och störd produktion av neurotransmitter-prekursorer (tryptofan, GABA). [B30], [B31]. Prækliniska studier visar att faikal mikrobiotatransplantation förbättrar ASD-liknande beteenden i musmodeller och modulerar serotonerga och glutamaterga signalvägar [B32], [B33].
Behov av adjuvant terapi
Trots intensiv forskning finns inga botande behandlingar för ASD:s kärnsymtom. Evidensbaserade interventioner inkluderar beteendeinriktade metoder (Applied Behavior Analysis, Early Start Denver Model) och symptomatisk farmakoterapi. [B34], [B35]. Risperidon och aripiprazol är de enda FDA-godkända läkemedlen för ASD, dock endast för irritabilitet och aggressiva beteenden, inte för kärnsymtom [B36], [B37]. Dessa antipsykotiska läkemedel är förknippade med betydande biverkningar (viktökning, metaboliskt syndrom, extrapyramidala symtom). [B38].
Komorbida symptom som ångest, sömnstörningar och sensorisk dysreglering behandlas ofta med bensodiazepiner, melatonin eller antidepressiva med begränsad evidens för effekt och säkerhet hos ASD-populationer. [B39], [B40]. Denna brist på säkra, effektiva terapier för komorbida symtom och bördan av polyfarmaci motiverar sökandet efter adjuvanta interventioner med få biverkningar [B41].
Eteriska oljor och deras huvudkomponenter (terpener) utgör en potentiell adjuvant terapioption, baserad på:
- prekliniska data om GABAerg modulering, serotonerg reglering, anti-neuroinflammatoriska och antioxidativa effekter;
- traditionell användning inom aromaterapi för ångest, sömn och humör;
- billiga säkerhetsprofilen vid korrekt användning [B42], [B43], [B44].
Neurobiologi av autismspektrumstörning
Denna rapport analyserar de neurobiologiska grunderna för ASD, de molekylära verkningsmekanismerna hos relevanta terpener, klinisk evidens hos ASD-populationer och utvecklar evidensbaserade doserings- och säkerhetsrekommendationer för användning av eteriska oljor som en del av en multimodal behandlingsstrategi.
GABA/Glutamat E/I-obalans
E/I-obalanshpotet är ett av de mest robusta neurobiologiska koncepten inom ASD-forskning. [B17], [B18]. Glutamat, den primära excitatoriska signalsubstansen, och GABA (γ-aminosmörsyra), den primära inhibitoriska signalsubstansen, reglerar neuronal excitabilitet, synaptisk plasticitet och nätverksdynamik. En obalans till förmån för excitatorisk transmission (ökad E/I-kvot) eller störd inhibitorisk kontroll kan leda till överexcitabilitet, störd sensorisk filtrering och nedsatt bearbetning av social information [B45], [B46].
Magnetresonansspektroskopi (MRS)-studier visar regionala förändringar: reducerade striatala glutamatkoncentrationer, förhöjda prefrontala GABA-nivåer i vissa studier och förändrade glutamat/GABA-förhållanden i sensoriska och associativa kortexområden [B19], [B20].
Postmortala analyser visar minskad uttrycksnivå av GAD65 och GAD67 (GABA-syntetiserande enzymer) i prefrontala cortex och cerebellum, samt förändrad densitet av GABAergiska interneuroner (specifikt parvalbumin-positiva interneuroner). [B47], [B48].
Det utvecklingsbiologiska perspektivet är kritiskt: GABA verkar excitatoriskt under tidiga utvecklingsstadier på grund av höga intracellulära kloridkoncentrationer (förmedlat av NKCC1-kotransportörer) och övergår postnatalt till hämmande effekt genom uttryck av KCC2-kotransportören [B49].
Fördröjd eller störd GABA-omkoppling har påvisats i ASD-musmodeller och korrigerats med bumetanid (NKCC1-hämmare), vilket ledde till förbättrat socialt beteende [B50], [B51].
En klinisk studie med bumetanid hos barn med ASD visade måttliga förbättringar i social kommunikation, men med metodologiska begränsningar [B52].
Glutamaterg dysfunktion manifesterar sig i förändrad NMDA- och AMPA-receptorsubtyps-expression, störd synaptisk plasticitet (long-term potentiation/depression) och överdriven glutamaterg transmission i specifika kretsar [B53], [B54].
Genetiska studier identifierade ASD-associerade mutationer i gener som kodar för glutamaterga synapser (GRIN2B, GRIA1, SLC1A1) [B55].
Prekliniska modeller visar att modulering av glutamaterg transmission (t.ex. med mGluR5-antagonister) kan förbättra ASD-liknande beteenden [B56].
Serotonin, dopamin och oxytocin
Monoaminerga system spelar centrala roller i social kognition, belöningsbearbetning och emotionell reglering – områden som är nedsatta vid ASD. [B57], [B58].
Serotonin (5-HT)
Hyperserotoninemi (förhöjda perifera serotoninnivåer) har påvisats hos 25–50 % av ASD-patienter, medan sambandet med centrala 5-HT-nivåer förblir oklart [B59], [B60]. Bildstudier visar nedsatt synteskapacitet för serotonin i frontala cortex och thalamus hos barn med ASD. [B61]. Genetiska varianter i serotonin-transportören (5-HTTLPR) och tryptofan-hydroxylas-2 (TPH2) är associerade med ASD-risk. [B62]. Prekliniska studier visar att serotonerg dysfunktion under kritiska utvecklingsfönster leder till ASD-liknande fenotyper [B63]. Selektiva serotoninåterupptagshämmare (SSRI) visar blandade resultat vid ASD: ingen effekt på kärnsymtom, måttliga effekter på repetitiva beteenden och oro [B64].
Dopamin (DA)
Dopaminerga mesolimbiska och mesokortikala kretsar medierar belöningsbearbetning, motivation och social förstärkning [B65]. ASD-patienter uppvisar minskad aktivering av den ventrala striatum vid sociala belöningar (ansikten, social interaktion) jämfört med icke-sociala belöningar. [B66], [B67]. PET-studien rapporterar ökad dopamintransportörs-densitet i striatum och reducerad D2-receptortillgänglighet [B68]. Genetiska studier identifierade ASD-associerade varianter i dopaminerga gener (DRD1, DRD2, DAT1). [B69]. Prekliniska modeller visar att dopaminerg dysfunktion leder till nedsatt social preferens och repetitiva beteenden [B70].
Oxytocin (OT)
Oxytocin är ett neuropeptid med en central roll i social bindning, tillit, social kognition och stressreglering [B71]. Patienter med ASD uppvisar reducerade plasmanivåer av oxytocin, förändrad expression av oxytocinreceptorn (OXTR) och genetiska varianter i OXTR och CD38 (oxytocinfrisättning). [B72], [B73]. Intranasal oxytocin förbättrade i vissa studier social kognition och ögonkontakt hos vuxna med ASD, men med inkonsekventa resultat och avsaknad av effekt i pediatriska kohorter. [B74], [B75]. Prekliniska studier visar att oxytocin reglerar GABAerg utveckling: Oxytocinadministrering under kritiska perioder korrigerade den fördröjda GABA-omkopplingen och förbättrade socialt beteende i ASD-musmodeller [B50], [B76]. Dessa fynd kopplar oxytonerga och GABAerga system mekanistiskt.
Neuroinflammation – Mikroglia, TNF-α och cytokiner
Kronisk neuroinflammation är en konvergent patofysiologisk mekanism vid ASD, stödd av postmortem-, bild- och biomarkörstudier [B77], [B78].
Aktivering av mikroglia
Mikroglia är de residenta immuncellerna i CNS och de reglerar synaptisk beskärning, neuronal utveckling och immunövervakning [B79]. Postmortala studier visar aktiverade mikroglia (ökad Iba1-expression, amöboid morfologi) i prefrontala cortex, lillhjärnan och vita substansen hos hjärnor med ASD. [B80], [B81]. PET-avbildning med translocatorprotein (TSPO)-ligander visade ökad mikrogliaaktivering i flera hjärnregioner hos levande ASD-patienter [B82]. Aktiverade mikroglia frisätter proinflammatoriska cytokiner, producerar ROS och kan fagocytera synaptiska strukturer, vilket leder till störd konnektivitet [B83].
Pro-inflammatoriska cytokiner
Förhöjda nivåer av TNF-α, IL-6, IL-1β och IFN-γ har påvisats i plasma, cerebrospinalvätska och post mortem hjärnvävnad från patienter med ASD [B84], [B85]. Modeller för maternal immunaktivering (MIA), där dräktiga möss behandlas med Poly(I:C) eller LPS, ger avkommor med ASD-liknande beteenden och förhöjda TNF-α-nivåer [B86], [B87]. TNF-α modulerar synaptisk transmission, minskar GABAerg inhibition och påverkar synaptisk plasticitet [B88]. Blockad av TNF-α i MIA-modeller förhindrade ASD-liknande fenotyper [B89].
Astrocytdysfunktion
Astrocyter reglerar glutamathomeostas (via glutamattransportörerna EAAT1/2), K+-buffring och metabolt stöd till neuroner [B90]. Postmortala studier visar astroglios (ökad GFAP-expression) i ASD-hjärnor [B91]. Dysfunktionella astrocyter kan bidra till överdriven ackumulering av extracellulärt glutamat och excitotoxicitet [B92].
Oxidativ stress och mitokondriell dysfunktion
Oxidativ stress uppstår som ett resultat av en obalans mellan ROS-produktion och antioxidativt försvar [B93]. Flera studier rapporterar ökade markörer för oxidativ stress vid ASD: ökad lipidperoxidation (malondialdehyd), protein-karbonylering, DNA-oxidation (8-OHdG) och minskad antioxidantkapacitet (glutation, superoxiddismutas, katalas) [B28], [B94], [B95].
Glutationnedbrytning
Glutation (GSH) är det primära intracellulära antioxidanten. ASD-patienter uppvisar reducerade GSH-nivåer i plasma och hjärna, ökad oxiderad glutation (GSSG) och reducerat GSH/GSSG-förhållande [B96], [B97]. Genetiska varianter i glutationssyntasenzymer (GCLC, GSS) är associerade med ASD-risk [B98].
Mitokondriell dysfunktion
5 till 30 % av ASD-patienter uppvisar biokemiska tecken på mitokondriell dysfunktion (förhöjt laktat, reducerad andningskedjeaktivitet) [B99], [B100]. Mitochondrierna är den huvudsakliga källan till ROS; mitokondriell dysfunktion leder till ökad ROS-produktion, minskad ATP-syntes och störd kalcium-homeostas [B101]. Prekliniska studier visar att mitokondriella antioxidanter (MitoQ) förbättrar ASD-liknande beteenden i musmodeller [B102].
Nrf2/HO-1-vägen
Den kärnrelaterade faktorn för erytroid 2 (Nrf2) är en huvudregulator av den antioxidativa responsen, som inducerar uttrycket av hemoxygenase-1 (HO-1), NAD(P)H:quinonoxidoreduktas-1 (NQO1), glutation-S-transferaser och andra antioxidantenzymer [B103]. Reducerad Nrf2-aktivitet har påvisats i ASD-modeller; Nrf2-aktivatorer (sulforafan) visade förbättringar i socialt beteende och kommunikation vid ASD i små kliniska studier [B104], [B105].
Tarm-hjärna-axeln
Den dubbelriktade kommunikationen mellan mag-tarmkanalen och centrala nervsystemet sker via neurala (Vagusnerven), endokrina (Hypotalamus-Hypofys-Binjure-axeln), immunologiska och metaboliska vägar. [B106]. ASD-patienter uppvisar en hög prevalens av gastrointestinala symtom (förstoppning, diarré, buksmärta) och dysbios. [B107], [B108].
Mikrobiom-Alterationen
Metaanalyser visar minskad mikrobiell diversitet vid ASD, ökat Firmicutes/Bacteroidetes-förhållande, minskat Bifidobacterium och Prevotella, ökat Clostridium och Desulfovibrio [B109], [B110]. Dessa förändringar korrelerar med symtomens svårighetsgrad [B111].
Mekanistiska kopplingar
Tarmmikrobiotan producerar signalsubstansföregångare (tryptofan → serotonin, glutamat → GABA), kortkedjiga fettsyror (butyrat, propionat, acetat) och modulerar immunförsvaret [B112]. Propionat-exponering inducerar ASD-liknande beteenden i gnagarmodeller [B113]. Fekal mikrobiotatransplantation (FMT) från ASD-patienter till bakteriefria möss överförde ASD-liknande beteenden och förändrade genuttryck i hjärnan (gener för serotonerga, glutamaterga synapser). [B32]. FMT från friska donatorer i ASD-musmodeller förbättrade socialt beteende och normaliserade nivåer av signalsubstanser [B114].
Vagusförmedling
Vagusnerven överför signaler från tarmen till hjärnstammen (Nucleus tractus solitarii) och vidare till limbiska och kortikala regioner [B115]. Präkliniska studier visar att elektroakupunktur förbättrade ASD-liknande beteenden hos VPA-musmodeller via vagusberoende mekanismer och mikrobiommodulation; vagotomi upphävde dessa effekter. [B116].
Synaptisk dysfunktion – Neuroliginer, SHANK-proteiner
Synaptisk dysfunktion är ett centralt patofysiologiskt koncept vid ASD, understött av genetiska, molekylära och elektrofysiologiska bevis. [B117], [B118].
Neuroligin-Neurexin-komplex
Neuroliginer (NLGN1-4) är postsynaptiska celladhesionsmolekyler som interagerar med presynaptiska neurexiner (NRXN1-3) och reglerar synaptisk differentiering, mognad och funktion [B119]. Mutationer i NLGN3 och NLGN4X har identifierats hos ASD-patienter [B120]. NLGN3-R451C-Knockin-möss uppvisar ökad hämmande transmission, störd social interaktion och repetitiva beteenden [B121]. NLGN4-knockoutmöss uppvisar nedsatt excitatorisk transmission och ASD-liknande fenotyper [B122].
SHANK-proteiner
SHANK1-3 är postsynaptiska skelettproteiner som organiserar glutamatreceptorer, aktincytoskelettet och signalmolekyler [B123]. SHANK3-mutationer är associerade med Phelan-McDermid-syndrom (22q13-deletion), vilket kännetecknas av autism, intellektuell funktionsnedsättning och språkdefekter. [B124]. SHANK3-utslagna möss uppvisar reducerad dendritisk ryggradsdensitet, störd synaptisk transmission och ASD-liknande beteenden [B125]. SHANK2-mutationer är också associerade med ASD; SHANK2-knockoutmöss uppvisar hyperaktivitet, repetitiva beteenden och nedsatt NMDA-receptorfunkton [B126].
Synaptisk plasticitet
Långtids potentiell (LTP) och långtids depression (LTD), cellulära korrelat till inlärning och minne, är störda i multipla musmodeller av ASD. [B127]. En förändrad balans av LTP/LTD kan bidra till störd erfarenhetsberoende mognad av nervbanor och försämrat socialt lärande. [B128].
Drabbade hjärnregioner
Strukturell och funktionell avbildning identifierar konsekventa förändringar i specifika hjärnregioner vid ASD [B129], [B130].
Prefrontala cortex (PFC)
PFC förmedlar exekutiva funktioner, arbetsminne, kognitiv flexibilitet och social kognition [B131]. ASD-patienter uppvisar reducerad PFC-aktivering under theory-of-mind-uppgifter, störd PFC-konnektivitet med bakre regioner och förändrad PFC-mikrostruktur [B132], [B133]. Postmortala studier visar ökad neuronaltäthet, reducerad minikolumnbredd och GABAerga brister i prefrontala cortex. [B134].
Amygdala
Amygdala är centralt för känslomässig bearbetning, social perception och rädsloinlärning [B135]. Barn med ASD visar tidig amygdala-förstoring (2–4 år), följt av normalisering eller volymreduktion i tonåren [B136]. Funktionella studier visar minskad amygdalaaktivering vid ansiktsbehandling och störd koppling mellan amygdala och PFC. [B137], [B138].
Lillhjärnan
Traditionellt kopplat till motorisk kontroll spelar lillhjärnan också roller i kognitiva processer, sensorisk förutsägelse och socialt lärande [B139].
Patienter med ASD uppvisar förlust av Purkinjeceller, minskad volym i lillhjärnan (särskilt vermis) och störd cerebello-kortikal konnektivitet [B140], [B141]. Cerebellär dysfunktion kan bidra till störd sensorisk prediktion, motoriska stereotypier och nedsatt förmåga att bearbeta social timing. [B142].
Hippocampus
Hippocampus förmedlar deklarativt minne, rumslig navigering och kontextuell bearbetning [B143]. ASD-patienter uppvisar förstoring av hippocampus under barndomen, förändrad hippocampus-konnektivitet och störd kontextuell rädsloinlärning [B144], [B145].
Striatum
Striatum (nucleus caudatus, putamen, nucleus accumbens) förmedlar belöningsbearbetning, vanemässigt lärande och motorisk kontroll [B146]. Patienter med ASD uppvisar striatumförstoring, minskad aktivering vid sociala belöningar och störd kortiko-striatal konnektivitet [B147], [B148].
Molekylära verkningsmekanismer för terpener vid ASD
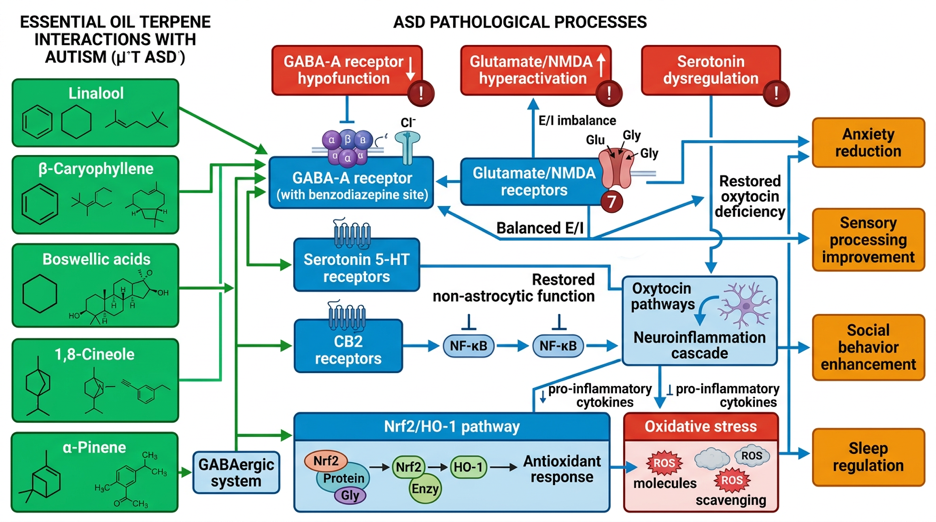
Figur 2: Terpenernas molekylära signalvägar vid autism-spektrumstörning – GABA/glutamatbalans, neuroinflammation och oxidativ stress
Terpener är en mangfaldig klass av naturprodukter som utgör huvudkomponenterna i eteriska oljor [B149]. Deras lipofila egenskaper möjliggör passage genom blod- hjärnbarriären och interaktion med neurala mål. [B150]. Det här avsnittet analyserar molekylära mekanismer för specifika terpener som angriper ASD-relaterade patofysiologiska mål.
Linalool – GABA-A/bensodiazepinreceptor, serotonin, antiinflammatorisk
Linalool (3,7-Dimetyl-1,6-oktadien-3-ol) är en monoterpenisk alkohol och en huvudkomponent i lavendel (Lavandula angustifolia, 25–45 %), koriander, bergamott och andra eteriska oljor [B151].
GABA-A-receptormodulering
Linalool uppvisar ångestdämpande effekter som blockeras av flumazenil (en bensodiazepinantagonist), vilket tyder på interaktion med bensodiazepinbindningsstället på GABA-A-receptorn. [B152]. I en modell för olfaktorisk exponering producerade linalool-ångor ångestdämpande effekter hos möss (Elevated Plus Maze, Light-Dark Box) som saknades hos anosmatiska möss (förstörelse av det olfaktoriska epiteliet), vilket demonstrerar en lukt-medierad effekt. [B152]. Elektrofysiologiska studier visar att linalool förstärker GABA-inducerade kloridströmmar i odlade neuroner [B153]. Denna GABAerga modulering är mycket relevant för ASD, eftersom GABAerga brister och E/I-obalans utgör centrala patofysiologiska mekanismer [B17], [B18].
Serotonerg modulation
Inhalation av linalool förändrade serotoninnivåer i råttors frontala cortex, vilket indikerar modulering av serotonerg transmission [B154]. I en studie med friska vuxna personer minskade 20 minuters inhalation av linalool depressivt humör och ökade parasympatisk aktivitet (ökad hjärtfrekvensvariabilitet). [B155]. Ex vivo-studier visar att linalool ökar intracellulära kalciumkoncentrationer i hypotalamiska oxytocinerga neuroner, vilket uppvisar en möjlig mekanism för oxytongerg modulering. [B155]. Dessa fynd kopplar linalool till serotonerga och oxytocinerga system, som är dysfunktionella vid ASD [B59], [B72].
Antiinflammatoriska effekter
Systematiska översikter rapporterar antiinflammatoriska egenskaper hos linalool i prekliniska modeller, inklusive minskning av TNF-α, IL-6, IL-1β och NF-κB-aktivering [B156]. I en stressinducerad modell av neuroinflammation minskade linalool mikrotrombocyaktivering och uttryck av proinflammatoriska cytokiner [B157]. Dessa anti-neuroinflammatoriska effekter är relevanta för ASD, eftersom kronisk neuroinflammation är en konvergent patofysiologisk mekanism [B77], [B78].
Farmakokinetik
Efter inhalativ exponering absorberas linalool snabbt och når systemcirkulationen; metaboliter (linalooloxider, glukuronider) påvisas i urin [B158]. Blod-hjärnbarriärpassagen är sannolikt möjlig på grund av lipofiliteten, men direkta data om farmakokinetik i centrala nervsystemet hos människa är begränsade. [B159].
β-Karyofyllen – CB2-agonist, NF-κB, neuroinflammation
β-Karyofyllen (BCP) är en sesquiterpenkolväte som finns i svartpeppar, kryddnejlika, cannabis (icke-psykoaktiv komponent), copaiba och andra växter [B160].
CB2-receptöragonism
BCP är en selektiv cannabinoid-typ-2-receptor (CB2)-agonist utan affinitet för CB1-receptorer, vilket möjliggör icke-psykoaktiva immunmodulerande effekter [B161]. CB2-receptorer uttrycks primärt på immunceller (mikroglia, perifera immunceller) och reglerar cytokinfrisättning, mikrogliaaktivering och neuroinflammation [B162]. Hos CB2-knockoutmöss saknades BCP-effekter, vilket bekräftar CB2-specificitet [B163].
Anti-neuroinflammatoriska effekter
BCP minskar neuroinflammation i flera prekliniska modeller: LPS-inducerad neuroinflammation, traumatisk hjärnskada, neurodegenerativa sjukdomar [B164], [B165]. Mekanismerna inkluderar minskad aktivering av mikroglia (reducerad Iba1-, CD11b-expression), suppression av proinflammatoriska cytokiner (TNF-α, IL-6, IL-1β) och modulering av NF-κB-signalvägar. [B166]. I en modell för maternal immunaktivering (MIA, ASD-relevant modell) förbättrade BCP-behandling ASD-liknande beteenden (sociala brister, repetitiva beteenden) och minskade neuroinflammation hos avkomman. [B167].
GABAerg och nitrinerg modulering
Farmakologiska studier av beteendet visar att ångestdämpande och antidepressiva effekter av BCP blockeras av flumazenil (bensodiazepinantagonist) och L-NAME (kväveoxidsyntashämmare), vilket indikerar inblandning av GABAerga och nitrinerga system [B168]. Dessa fynd antyder att BCP indirekt kan modulera GABAerg funktion genom CB2-medierad minskning av neuroinflammation.
Antioxidativa effekter
BCP uppvisar antioxidativa egenskaper genom reduktion av ROS, lipidperoxidation och ökning av endogena antioxidant-enzymer (SOD, katalas, GSH) [B169]. I en modell för oxidativ stress skyddade BCP neuroner från H₂O₂-inducerad apoptos [B170].
Klinisk relevans för AST
Kombinationen av CB2-medierad minskning av neuroinflammation, antioxidativa effekter och indirekt GABAerg modulation gör BCP till en mekanistiskt plausibel kandidat för ASD-adjuvant terapi, särskilt hos patienter med neuroinflammatoriska signaturer. [B171].
Boswelliasyror – Nrf2/HO-1, anti-neuroinflammatoriska
Boswelliasyror är pentacykliska triterpensyror från kådan från Boswellia-arter (skrän), inklusive β-boswelliasyra, 11-keto-β-boswelliasyra (KBA) och 3-O-acetyl-11-keto-β-boswelliasyra (AKBA) [B172].
Nrf2/HO-1-aktivering
Boswelliasyror aktiverar Nrf2-signalvägen, en masterregulator för det antioxidativa svaret [B173]. I en murin neuroinflammatorisk modell ökade behandling med boswelliaextrakt Nrf2-kärnlokalisering och HO-1-uttryck i hippocampus och cortex [B174]. HO-1 katalyserar nedbrytning av hem till biliverdin (antioxidativt), kolmonoxid (antiinflammatoriskt, kärlvidgande) och järn; HO-1-induktion skyddar mot oxidativ stress och neuroinflammation [B175].
Anti-inflammatoriska mekanismer
Boswelliasyror minskar mikroglia- och astrocytaaktivering (reducerad Iba1-, GFAP-expression), dämpar proinflammatoriska cytokiner (TNF-α, IL-6, IL-1β) och hämmar NF-κB- och MAPK-signalvägar [B176], [B177]. I en LPS-inducerad modell av neuroinflammation förbättrade Boswellia-behandling kognitiva brister och minskade markörer för neuroinflammation [B178].
5-Lipoxygenas (5-LOX)-hämning
Boswelliasyror, särskilt AKBA, hämmar 5-LOX, ett enzym som metaboliserar arakidonsyra till proinflammatoriska leukotriener [B179]. 5-LOX-hämning minskar leukotrienmedierad inflammation och har i prekliniska studier associerats med neuroprotektiva effekter [B180]. Dock är bevisen för 5-LOX-hämning som den primära mekanismen för Boswellia's neuroprotektion begränsade i den befintliga litteraturen; Nrf2/HO-1-aktivering verkar vara den dominerande mekanismen [B174].
Preklinisk evidens vid ASD-relevanta modeller
I en ASD-musmodell inducerad av valproinsyra (VPA) förbättrade behandling med boswelliaextrakt social interaktion, minskade repetitiva beteenden och normaliserade markörer för neuroinflammation [B181]. Dessa fynd stöder hypotesen att Boswellia-förmedlad minskning av neuroinflammation och antioxidativa effekter kan förbättra ASD-relaterade beteenden.
1,8-Cineol – GABA-A/bensodiazepin, ångestdämpande
1,8-Cineol (eukalyptol) är en monoterpenoxid och huvudkomponent i eukalyptus (Eucalyptus globulus, 70–90 %), rosmarin, tea tree och lagerblad. [B182].
GABA-A-receptormodulering
Inhalation av 1,8-cineol gav dosberoende anxiolytiska och antidepressiva effekter hos möss (Elevated Plus Maze, Forced Swim Test, Tail Suspension Test). [B183]. Dessa effekter blockerades av flumazenil, vilket indikerar en interaktion med bensodiazepinbindningsstället på GABA-A-receptorn. [B183]. Doseringsberoende demonstrerades: låga doser (0,1–1 µg/mus, inhalativt) visade anxiolytiska effekter, medan högre doser (10 µg/mus) gav sedativa effekter [B184].
Antiinflammatoriska och antioxidativa effekter
1,8-Cineol uppvisar antiinflammatoriska egenskaper genom hämning av NF-κB, minskning av TNF-α, IL-1β och leukotrienproduktion [B185]. I en LPS-inducerad modell av neuroinflammation minskade 1,8-cineol mikrogliapåverkan och oxidativ stress. [B186].
Säkerhetsaspekter
1,8-Cineol är kontraindicerat för barn under 3 år på grund av risken för struplockskramp och andningsdepression [B187]. Vid korrekt användning (inhalativ, låga doser) visar 1,8-cineol en gynnsam säkerhetsprofil hos vuxna och äldre barn [B188].
α-Pinen – GABAerg
α-Pinen är en cyklisk monoterpen som finns i tall, rosmarin, salvia och cannabis [B189].
GABAerg modulering
Systematiska översikter associerar α-pinen med modulering av GABAerg neurotransmission, trots att direkta mekanistiska studier är begränsade [B190]. I beteendemodellen visar α-pinen ångestdämpande och sederande effekter, vilket indikerar GABAerga mekanismer. [B191]. Elektrofysiologiska data om direkt interaktion med GABA-A-receptorer saknas i den befintliga litteraturen.
Antiinflammatoriska och neuroprotektiva effekter
α-Pinen uppvisar antiinflammatoriska egenskaper genom hämning av NF-κB och reduktion av proinflammatoriska cytokiner [B192]. I en ischemisk reperfusionsmodell skyddade α-pinen mot neuronal skada och minskade oxidativ stress [B193].
Acetylkolinesteras (AChE)-hämning
Vissa studier rapporterar AChE-inhiberande aktivitet från α-pinen, vilket skulle kunna förstärka kolinerg neurotransmission [B194]. Men relevansen för ASD är oklar, eftersom kolinerg dysfunktion inte är en primär patofysiologisk mekanism vid ASD. [B195].
Ylang-Ylang/Cananga odorata – Serotonin/Dopamin i VPA-modell
Ylang-ylang (Cananga odorata) eterisk olja innehåller linalool, geraniol, β-karyofyllen och andra terpener [B196].
Preklinisk evidens i VPA-ASD-modellen
Inhalation av eterisk olja från Cananga odorata förbättrade ångestliknande beteenden, social interaktion och kognitiv funktion hos råttor med valproinsyreinducerad ASD [B197]. Neurokemiska analyser visade att Cananga odorata modulerade serotonin- och dopaminmetabolismen i prefrontala cortex och hippocampus: förhöjda serotoninnivåer, reducerade 5-HIAA/serotonin-förhållanden (vilket indikerar minskad serotoninomfattning) och normaliserade dopaminnivåer [B197].
Mekanistisk tolkning
Förbättring av ASD-liknande beteenden genom modulering av serotonerga och dopaminerga system är mekanistiskt plausibel, eftersom båda systemen är dysfunktionella vid ASD [B57], [B58]. De specifika komponenterna av Cananga odorata som är ansvariga för dessa effekter har inte identifierats; linalool och β-karyofyllen är troliga kandidater baserat på deras kända mekanismer [B152], [B161].
Begränsningar
Dessa fynd härrör från en enda preklinisk modell (VPA-råttor); replikering i andra ASD-modeller och translation till humana studier saknas [B197].
Specifika eteriska oljor och kliniska bevis vid ASD
Lavendel/Linalool – ABA-överensstämmelse, Oxytocin-neuronen
Kliniska bevis vid ASD
En liten pilotstudie (n = 12 barn med ASD) undersökte lavendel-aromaterapi som ett komplement under Applied Behavior Analysis (ABA)-terapi. [B198]. Lavendel diffunderades som “20 droppar i 100 ml” (förmodligen vatten eller bärarolja, detaljer oklara) under terapisessioner. Studien rapporterade högre följsamhetspoäng (samarbete, uppgiftsutförande) under lavendel-exponering jämfört med kontrollsessioner. [B198]. Begränsningarna inkluderar mycket liten stickprovsstorlek, brist på randomisering, oklar blindning och bristande standardiserad dosering.
En observationsstudie med fyra barn med ASD och svåra inlärningssvårigheter undersökte aromaterapimassage (lavendel, geranium, mandarin i basolja) integrerat i dagliga rutiner [B199]. Studien rapporterade ökade beteenden relaterade till delad uppmärksamhet (blickkontakt, gemensamma aktiviteter) efter introduktionen av aromaterapimassage. [B199]. Begränsningarna inkluderar bristande kontroll, liten urvalsstorlek och förväxling från massageeffekter (taktil stimulering, social interaktion).
Mekanistisk grund
De kliniska fynden överensstämmer med prekliniska mekanismer: linalool-medierad GABAsyntetisk modulering kan minska ångest och förbättra samarbete; serotonerg och oxytocinerg modulering kan främja social uppmärksamhet och interaktion. [B152], [B155]. En studie med friska vuxna visade att lavendelinhalation minskade depressivt humör och i ex vivo-experiment ökade intracellulära kalciumkoncentrationer i hypotalamiska oxytocineroner. [B155]. Denna oxytonerga modulation är mycket relevant för ASD, eftersom oxytocin-dysfunktion är en etablerad patofysiologisk mekanism [B72].
Bevisbegränsningen
Den kliniska evidensen för lavendel vid ASD är av mycket låg kvalitet: små urval, bristande randomisering, otillräcklig blindning, heterogena interventioner och bristande replikering [B198], [B199]. Högkvalitativa, randomiserade kontrollerade studier med standardiserade lavendelpreparat, objektiva utfallsmått och tillräcklig styrka saknas.
Bergamott – RCT vid ASD-barn, resultat ej signifikant
Randomiserad kontrollerad studie
En randomiserad, dubbelblind studie undersökte eterisk olja från bergamott (Citrus bergamia) vid medicinskt inducerad ångest hos barn med ASD (n = 42). [B200]. Barn randomiserades till 15 minuters inhalation av bergamottolja eller placebo (vatten) före medicinska undersökningar. Primärt utfallsmått var ångest mätt med Modified Yale Preoperative Anxiety Scale (mYPAS). Efter justering för ångest vid baslinjen visade bergamottgruppen ingen signifikant minskning av ångest jämfört med placebo (p > 0.05) [B200].
tolkning
Detta är den enda identifierade randomiserade kontrollerade studien med eteriska oljor hos barn med ASD. Det negativa resultatet kan ha flera orsaker: (1) kort exponeringstid (15 minuter) kan vara otillräcklig; (2) akut ångest i medicinska miljöer kan vara svårare att modulera än kronisk ångest; (3) individuell variabilitet i luktkänslighet och preferenser; (4) potentiella takeffekter vid redan hög baslinjeångest [B200].
Präkliniska mekanismer
Prekliniska studier visar ångestdämpande effekter av bergamott i gnagarmodeller [B201]. Intressant nog blockerades dessa effekter inte av flumazenil, vilket tyder på icke-bensodiazepin-medierade mekanismer och skiljer bergamott från linalool och 1,8-cineol [B201]. Bergamott innehåller linalool, linalylacetat, limonen och bergapten; de specifika komponenterna som är ansvariga för ångestdämpande effekter är oklara [B202].
Kliniska implikationer
Det negativa resultatet av denna RCT dämpar förväntningarna på bergamott vid akut ångest hos ASD. Ytterligare studier med längre exponeringstider, kroniska ångestutfall och andra ASD-populationer är nödvändiga innan bergamott kan rekommenderas för ASD. [B200].
Frankincense / Boswellia – Anti-neuroinflammatorisk, Nrf2
Kliniska bevis vid ASD
Det identifierades inga randomiserade kontrollerade studier med eterisk olja av rökelse (Boswellia) eller Boswellia-extrakt hos ASD-patienter. Föreliggande litteratur saknar ASD-specifik klinisk data för rökelse. [B203].
Preklinisk evidens
Som beskrivs i avsnitt 3.3 uppvisar boswelliasyror robusta anti-neuroinflammatoriska och antioxidativa effekter i prekliniska modeller, inklusive Nrf2/HO-1-aktivering, mikroglia-hämning och förbättring av ASD-liknande beteenden i VPA-musemodeller. [B174], [B181]. Denna mekanistiska rimlighet stöder hypotesen att Boswellia kan vara användbart hos ASD-patienter med neuroinflammatoriska signaturer.
Klinisk tillämpning av andra indikationer
Boswellia-extrakter (orala) används kliniskt för inflammatoriska sjukdomar (artrit, astma, inflammatoriska tarmsjukdomar), med måttliga bevis för effektivitet och en gynnsam säkerhetsprofil [B204]. Inhalativ användning av rökelse eterisk olja är traditionell, men kliniska farmakokinetiska och effektdata saknas [B205].
Forskningsbehov
Kontrollerade kliniska studier med Boswellia (oralt eller inhalativt) hos patienter med ASD, helst stratifierade efter biomarkörer för neuroinflammation, är nödvändiga för att utvärdera klinisk effekt. [B206].
Cederträ/Zeder – Sedering, GABAerg
Kliniska och prekliniska bevis
Den föreliggande litteraturen innehåller inga specifika mekanistiska eller kliniska data för eterisk olja från cederträ (Cedrus atlantica, Juniperus virginiana) vid ASD eller i ASD-relevanta prekliniska modeller [B207]. Traditionella användningsområden inkluderar sedering och ångestreducering, men det saknas vetenskapliga belägg för GABAerga mekanismer eller klinisk effekt i den analyserade litteraturen [B208].
Huvudkomponenter
Cederolje inneholder cedrol (sesquiterpen alkohol), α-cedren og thujopsen [B209]. Cedrol har associerats med lugnande egenskaper, men direkta studier av interaktion med GABA-A-receptorer eller modulering av neuroinflammation saknas [B210].
Slutsats
Det finns otillräcklig evidens för ceder i samband med ASD; rekommendationer kan inte göras baserat på den föreliggande litteraturen [B207].
Vetiver – Uppmärksamhet
Kliniska och prekliniska bevis
Den föreliggande litteraturen innehåller inga specifika data för eterisk olja av vetiver (Vetiveria zizanioides) vid ASD eller i ASD-relevanta modeller [B211]. Anekdotiska rapporter och traditionella användningsområden antyder effekter på uppmärksamhet och hyperaktivitet, men kontrollerade studier saknas [B212].
Huvudkomponenter
Vetiverolja innehåller vetiverol, khusimol, α-vetivon och β-vetivon (sesquiterpeniska alkoholer och ketoner). [B213]. Mekanistiska studier av dessa komponenter i förhållande till signalsubstanssystem eller neuroinflammation saknas i den analyserade litteraturen [B214].
Slutsats
Bevisen för vetiver vid ASD är otillräckliga; ytterligare forskning behövs innan rekommendationer kan göras [B211].
Ylang-Ylang – Serotonin/Dopamin, VPA-modell
Preklinisk evidens
Som beskrivs i avsnitt 3.5, förbättrade inhalation av eterisk olja från Cananga odorata (Ylang-Ylang) ASD-liknande beteenden hos VPA-exponerade råttor och modulerade serotonin- och dopaminmetabolismen i prefrontala cortex och hippocampus. [B197]. Detta är den enda identifierade studien med Ylang-Ylang i en ASD-relaterad modell.
Kliniska bevis vid ASD
Inga kliniska studier med ylang-ylang hos ASD-patienter har identifierats [B215].
Klinisk tillämpning av andra indikationer
Ylang ylang används traditionellt mot ångest, humör och avslappning. En liten studie med friska vuxna visade att inandning av ylang ylang sänkte blodtrycket och hjärtfrekvensen och ökade det subjektiva lugnet [B216]. Mekanistiska studier av serotonerga och dopaminerga effekter hos människor saknas [B217].
Slutsats
Den prekliniska evidensen är lovande, men translation till studier på människor med ASD saknas. Ylang-Ylang skulle kunna övervägas som en kandidat för kliniska studier vid ASD, särskilt för ångest- och stämningssymtom. [B197].
Ytterligare indicerade eteriska oljor vid ASD
Baserat på aktuell litteraturforskning identifierades följande ytterligare eteriska oljor som kan vara relevanta för ASD via neurobiologiskt rimliga mekanismer.
Ingefära (Zingiber officinale) – AKT/GSK3β-signalväg
Aktiva ingredienser
6-Gingerol
6-Shogaol
Zingiberen
β-Sesquiphellandren
Receptorer och verkningsmekanismer
– AKT/GSK3β-signalväg: Ökad fosforylering → förbättrad synaptisk plasticitet
– Anti-neuroinflammatoriskt: Minskning av STAT3-fosforylering och TNF-α
Neurogenes: Förbättrade markörer för hippocampal neurogenes i prekliniska modeller
– Gliosämpning: Minskad reaktiv astroglios
Kliniska/prekliniska bevis
- Präklinisk studie (VPA-musmodell)
– Orala ingefäraextrakt (4 veckor, från 6:e levnadsveckan) förbättrade signifikant social interaktion, minskade ångestliknande beteende och förbättrade minnesprestation [B347]
Mekaniskt: Uppreglering av AKT/GSK3β, hämning av STAT3/TNF-α, förbättrad synapsbildning
– Kliniska RCT:er för ASD saknas fortfarande, men prekliniska bevis är starka och mekanistiskt väl underbyggda
Dosering (Organik Aromas Nebulizer 3.0)
– 2–3 droppar för 15–25 m²
– Kombination med lavendel (1:2) för avslappnande effekt
– Användning: 30 min före sociala aktiviteter
Säkerhet
Väl tolererat; använd med försiktighet till barn < 3 år.
Sandelträ (Santalum album) – α/β-santalol
Aktiva ingredienser
α-Santalol (50–55%)
β-Santalol (20–25%)
Sesquiterpen
Receptorer och verkningsmekanismer
– Serotonerg modulation: α-Santalol modulerar 5-HT-receptorer (prekliniskt)
– Ångestdämpande: GABAerg aktivitet dokumenterad i djurmodeller
– Lugnande/Avslappnande: Lämplig för sensorisk överkänslighet vid AST
Klinisk evidens
– Sandelträ var en del av Hiwa-sirapens RCT (persisk medicin, flera örter) hos barn med ASD, effekter kan inte isoleras och tillskrivas. [B348]
– Enskilda studier av anxiolys vid friska försöksdeltagare visar lugnande effekt
– Direkta ASD-specifika studier med rent sandelträolja saknas
Dosering (Organik Aromas Nebulizer 3.0)
– 3–4 droppar för rum på 15 till 25 m²
– Särskilt lämplig för kvälls-/sömndrituale för barn med ASD som har sömnsvårigheter
– Rekommenderas i kombination med lavendel (1:1)
Säkerhet
Mycket bra tolererat; en av de säkraste eteriska oljorna för barn (från 2 års ålder).
Terpenblandningar (inspirerade av cannabis) – Prosociala effekter
Aktiva ingredienser
Myrcen
Limefrukter
Linalool
Tall
β-Caryophyllen (som blandning)
Receptorer och verkningsmekanismer
– CB1/CB2-agonism: Terpenblandningar förstärker CBD-effekter (entourageeffekt)
– GABAerg modulation: Linalool + Myrcen synergistiskt
– Prosocial effekt oberoende av CBD påvisad
Klinisk evidens
- BTBR-musmodell (ASD-modell)
Inhalerade terpenblandningar (myrcen, limonen, linalool, pinen, β-karyofyllen) visade akuta prosociala effekter oberoende av CBD [B349] – Terpenblandningar förstärkte CBD:s effekt vid kombinerad administrering
– Hänvisning till entourageeffekten som behandlingsprincip för ASD
Dosering (Organik Aromas Nebulizer 3.0)
– Blandningsrekommendation: Lavendel (2 dl) + Svartpeppar (1 dl) + Bergamott (1 dl) + Rosmarin (1 dl)
– Rum 15 är 25 m²: 5 droppar totalblandning
– Användning: 30 minuter före sociala aktiviteter eller gruppterapi
Dosrekommendationer för Organik Aromas Nebulizer 3.0
Der Organik Aromas Nebulizer 3.0 använder Venturi-principen (Bernoulli-effekten) för vattenfri kallförstoftning av eteriska oljor [B218]. Till skillnad från ultraljuds- eller värmediffusorer, som späder ut eller värmer upp oljor med vatten, producerar nebulisatorn fina aerosolpartiklar (1 till 5 µm) av rena eteriska oljor utan termisk nedbrytning [B219]. Detta möjliggör högre koncentrationer av flyktiga terpener i luften och potentiellt starkare farmakologiska effekter [B220].
Venturiprincipen och kallförstoftning
Venturi-principen beskriver tryckminskningen i en vätska när den strömmar genom en förträngning [B221]. I nebulisatorn försätts tryckluft genom en munstycke, som skapar ett undertryck som suger upp eterisk olja ur ett behållare och finfördelar den till små droppar. [B222]. Dessa droppar frigörs ut i inomhusluften, där de andas in eller sedimenterar på ytor [B223].
Fördelar med kallförstoftning
- Bevarande av termolabila komponenter (ingen värmenedbrytning) [B224]
– Högre koncentration av flyktiga terpener (ingen vattenutspädningseffekt) [B225]
– Fin partikelstorlek (1 till 5 µm) möjliggör djup penetration i luftvägarna [B226]
Nackdelar
– Högre kostnader jämfört med ultraljudsdiffusorer [B227]
– Högre oljeförbrukning [B228]
– Överdosiseringsrisk vid felaktig användning [B229]
Volymspecifika droppmängder
Doseringsrekommendationerna baseras på rumsvolym, luftväxlingshastighet, önskad terpenkoncentration och exponeringstid. Följande rekommendationer är konservativa och tar hänsyn till ASD-specifik sensorisk överkänslighet [B230].
Tabell 1: Dos rekommendationer för Organik Aromas Nebulizer 3.0 beroende på rumsstorlek

Anteckningar
- Droppvolym: 1 droppe ≈ 0,05 ml eterisk olja [B231]
- Startdos: Börja med det lägsta rekommenderade droppantalet och öka gradvis över 1 till 2 veckor för att bedöma toleransen [B232]
- Individuell anpassning ASD-patienter med uttalad överkänslighet för lukter kan behöva lägre doser eller längre pauser [B233]
- Ventilation: God ventilation är väsentlig för att undvika överdriven ackumulering [B234]
Användningstid och pauser
Användningstid
15 till 30 minuter per session är tillräckliga för att uppnå terapeutiska terpenkoncentrationer, baserat på prekliniska inhalationsstudier [B183], [B184]. Längre användning (>30 minuter) ökar risken för sensorisk överbelastning och olfaktorisk anpassning (reducerad uppfattning vid kontinuerlig exponering) [B235].
Paus
30 till 60 minuters pauser mellan användningar möjliggör olfaktorisk återhämtning, metabolism av inhalerade terpener och minskning av koncentrationer i rumsluften [B236]. Kontinuerlig exponering utan pauser kan leda till huvudvärk, illamående och sensorisk aversion, särskilt hos patienter med autism (ASD) som har sensorisk överkänslighet. [B237].
Dagliga applikationer
2 till 3 appliceringar per dag (morgon, eftermiddag, kväll) räcker för de flesta indikationer [B238]. Fler än 4 applikationer per dag ökar risken för biverkningar utan ytterligare terapeutisk nytta [B239].
Tidsspecifika protokoll
Morgon (aktiverande, fokuserande)
- Olja Rosmarin (1,8-Cineol, α-Pinen), pepparmynta (mentol, menton), citron (limonen), bergamott (linalool, linalylacetat)
- Dosering: 3–5 droppar för 20–30 m² rum
- Varaktighet: 15–20 minuter
- Mål: Främjande av vakenhet, uppmärksamhet och kognitiv funktion för skola/terapi [B240]
Eftermiddag (balanserande, stämningsreglerande)
- Olja Bergamott, Ylang-ylang, Rosengeranium, Rökelse
- Dosering: 3–5 droppar för 20–30 m² rum
- Varaktighet: 20–30 minuter
- Mål: Minskning av eftermiddagsångest, stämningsstabilisering, övergång mellan aktiviteter [B241]
Kväll (lugnande, sömngivande)
- Olja Lavendel (Linalool, Linalylacetat), Rysk kamomill (Estrar), Cederträ (Cedrol), Vetiver
- Dosering: 4–6 droppar för 20–30 m² rum
- Varaktighet: 30 minuter före sänggåendet
- Mål: Reduktion av kvällsångest, främjande av avslappning och insomning [B242]
ASD-specifika särdrag
Sensorisk överkänslighet 60–90 % av ASD-patienter uppvisar överkänslighet för lukter eller motvilja mot vissa dofter [B243]. En gradvis introduktion är väsentlig:
- Vecka 1: Exponering utanför rummet (olja diffunderar in i angränsande rum) för att bedöma acceptans [B244]
- Vecka 2: Lägsta dos (2 droppar) i 10 minuter, patienten utanför rummet under nebulisering [B245]
- Vecka 3: Lägsta dos, patient i rummet, observation av tecken på tolerans (avslappning, ingen aversion) [B246]
- Vecka 4+ Gradvis ökning av dos och varaktighet baserat på individuell tolerans [B247]
Preferensbaserat urval
Låt patienten (om möjligt) lukta på olika oljor och uttrycka preferenser. Olfaktorisk preferens korrelerar med terapeutisk acceptans och följsamhet. [B248].
Visuella och auditiva ledtrådar
Använd visuella timers eller ljudsignaler för att visa början och slutet av aromaterapisessionen, vilket ökar förutsägbarhet och kontroll för personer med autism. [B249].
Föräldra-/vårdnadshavarträning
Utbilda föräldrar och vårdgivare i att observera tecken på tolerans (avslappning, förbättrad uppmärksamhet) och aversion (grimas, tillbakadragande, agitation). [B250].
Kvalitetskrav – GC/MS-analys
Gaskromatografi-masspektrometri (GC/MS)
GC/MS är guldstandarden för kemisk karakterisering av eteriska oljor [B251]. GC separerar flyktiga komponenter baserat på kokpunkt och polaritet; MS identifierar komponenter baserat på massa-till-laddning-förhållande [B252]. GC/MS-analys möjliggör:
- Identifiering av huvudkomponenter: Verifiering av närvaron av terapeutiskt relevanta terpener (t.ex. linalool i lavendel, 1,8-cineol i eukalyptus) [B253]
- Kvantifiering Procentuell sammansättning av varje komponent [B254]
- Detektion av förfalskningar: Identifiering av syntetiska tillsatser, utspädningsmedel eller föroreningar [B255]
- Batch-till-batch-konsistens Säkerställande av reproducerbar sammansättning [B256]
Kvalitetskriterier för terapeutiska eteriska oljor
- 100 % ren
Inga syntetiska tillsatser, bäroljor eller förtunningsmedel [B257] - Terapeutisk kvalitet
Oljor från växter som odlats, skördats och destillerats under optimala förhållanden [B258] - Organisch/Wildsammlung
Fri från bekämpningsmedel, herbicider och kemiska gödningsmedel [B259] - GC/MS-certifikat
Varje laddning bör levereras med analyscertifikat för GC/MS som dokumenterar huvudkomponenter och renhet [B260] - Botaniska namnet
Entydig identifiering av växtart (t.ex. Lavandula angustifolia, inte bara “lavendel”) [B261] - Ursprungsland och destillationsdatum
Transparens om ursprung och färskhet [B262]
Lagerhantering
Eteriska oljor bör förvaras i mörka glasflaskor vid sval temperatur (<25°C) för att minimera oxidation och nedbrytning [B263]. Oxiderade oljor kan orsaka hudirritation och uppvisa reducerad terapeutisk effekt [B264].
Säkerhetsanvisningar och kontraindikationer
ASD-specifik sensorisk överkänslighet
Sensoriska bearbetningssvårigheter finns hos 60–90 % av ASD-patienterna och manifesteras som hypersensitivitet (överstimulering) eller hyposensitivitet (understimulering) gentemot sensoriska stimuli [B265]. Olfaktorisk överkänslighet kan leda till aversion, ångest, illamående och undvikande beteende [B266].
Riskhantering
- Kandidatintroduktion Som beskrivs i avsnitt 5.4, börja med minimala doser utanför patientrummet [B244]
- Preferensscreening Testa oljor före terapeutisk användning för att identifiera aversiva reaktioner [B248]
- Avbrottskriterier: Sluta omedelbart vid tecken på lidande (agitation, tillbakadragande, illamående, huvudvärk) [B267]
- Alternativa applikationsvägar: Vid inhalationsintolerans, överväg topisk applicering (spädd i bärarolja, lapptest krävs) eller passiv diffusion (olja på bomullsrondell i rumshörn). [B268]
Epilepsirisk – kamferhaltiga oljor
10–30 % av ASD-patienter har komorbid epilepsi [B269]. Vissa eteriska oljor, särskilt de som innehåller kamfer, kan sänka kramptröskeln och utlösa anfall [B270].
Kontraindicerade oljor vid epilepsi
- Kamfer (Cinnamomum camphora): Innehåller kamfer (upp till 50 %), ett känt krampfmedel [B271]
- Rosmarin (Rosmarinus officinalis, Kamferkemotyp): Innehåller 15–25 % Kamfer [B272]
- Salvia (Salvia officinalis): Innehåller tujon (10–40 %), en GABA-A-antagonist med kramplösande egenskaper [B273]
- Ysop (Hyssopus officinalis): Innehåller pinocamfon och isopinocamfon, som kan orsaka anfall [B274]
- Fänkål (Foeniculum vulgare): Innehåller trans-anetol, som i höga doser kan verka kramplikt [B275]
Säkra alternativ vid epilepsi: – Lavendel (Lavandula angustifolia, Linalool-Chemotyp): Antikonvulsiva egenskaper i prekliniska studier [B276] Romersk kamomill (Anthemis nobilis): Inga kända prokonvulsiva effekter [B277] – Bergamott (Citrus bergamia, bergaptenfri): Säker vid epilepsi [B278]
Klinisk rekommendation: An ASD-patienter med epilepsi bör endast oljor utan kamferhaltiga eller tujonhaltiga komponenter användas. Konsultation med en neurolog före aromaterapi är rekommenderat. [B279].
Eukalyptus hos barn under 3 år
1,8-Cineol (huvudkomponent i eukalyptus) är kontraindicerat hos barn under 3 år på grund av risken för laryngospasm, bronkospasm och andningsdepression [B280]. Mekanismerna inkluderar direkt irritation av luftvägarnas slemhinna och utlösning av vagala reflexer [B281].
Oljer som är kontraindicerade för barn <3 år
– Eukalyptus (Eucalyptus globulus, E. radiata): 70–90 % 1,8-Cineol [B282]
– Rosmarin (Rosmarinus officinalis, 1,8-Cineol-kemotyp): 40–50 % 1,8-Cineol [B283]
– Tea tree (Melaleuca alternifolia): 5–15 % 1,8-Cineol (lägre risk, men försiktighet bör iakttas) [B284]
– Pepparmynta (Mentha piperita): Mentol kan orsaka liknande irritationer i luftvägarna [B285]
Säkra alternativ för barn <3 år
– Lavendel (Lavandula angustifolia): Säkert från födseln vid korrekt spädning [B286]
– Romersk kamomill (Anthemis nobilis): Säker från födseln [B287]
– Mandarin (Citrus reticulata): Säkert från 6 månader [B288]
Klinisk rekommendation
Undvik oljor rika på 1,8-cineol hos barn med ASD under 3 år.
Hos barn 3–6 år används låga doser (50 % av vuxendosen) och de ska observeras noga för luftvägssymtom [B289].
Interaktioner med antipsykotika: Risperidon, Aripiprazol
Risperidon och aripiprazol är de enda FDA-godkända läkemedlen för irritabilitet vid AST [B290]. Båda metaboliseras via cytokrom P450-enzymer (CYP2D6, CYP3A4) [B291]. Vissa eteriska oljor och terpener kan hämma eller inducera CYP-enzymer, vilket kan leda till förändrade läkemedelsnivåer [B292].
Potentiella interaktioner:
Bergamott
Innehåller bergapten (furanokoumarin), som hämmar CYP3A4 [B293]. Teoretiskt sett skulle bergamott kunna öka nivåerna av risperidon och aripiprazol, vilket kan förstärka biverkningar (sedering, extrapyramidala symtom). [B294]. Men den inhalativa exponeringen är betydligt lägre än det orala intaget (grapefruktjuiceeffekt); den kliniska relevansen av inhalativ bergamott-exponering är oklar. [B295].
Rekommendation
Använd bergamottfri bergamott (FCF, furanokumarinfri) hos patienter som behandlas med risperidon/aripiprazol [B296].
Grapefrukt
Grapefrukt eterisk olja innehåller furanokumariner och hämmar CYP3A4 [B297]. Undvik hos patienter som tar CYP3A4-metaboliserade läkemedel [B298].
Johannesört (Hypericum perforatum)
Även om det inte är en typisk eterisk olja, nämns Johannesört ibland i aromaterapikontexter. Johannesört är en potent CYP3A4-inducerare och kan minska nivåerna av risperidon/aripiprazol, vilket leder till minskad effekt [B299]
Kontraindikation Undvik Johannesört hos patienter som tar antipsykotika [B300].
Lavendel, Kamomill, Cederträ
Inga kända kliniskt relevanta CYP-interaktioner [B301]. Säkert hos patienter som får risperidon/aripiprazol [B302].
Farmakodynamiska interaktioner
Sederande eteriska oljor (lavendel, kamomill, cederträ) kan ha adderande sederande effekter med antipsykotika [B303]. Detta kan vara terapeutiskt önskvärt (t.ex. sömninducerande) men kräver övervakning på överdriven sedering [B304].
Klinisk rekommendation
Informera förskrivande läkare om aromaterapi-tillämpning. Använd bergaptenfria oljor hos patienter som behandlas med CYP3A4-substrat. Övervaka förändrade läkemedels-effekter (sedering, biverkningar). [B305].
Kvalitetskrav: GC/MS-analys
Som beskrivs i avsnitt 5.5 är GC/MS-analys avgörande för att säkerställa renhet, sammansättning och frånvaro av föroreningar [B251]. Förfalskade eller förorenade oljor kan orsaka allergier, hudirritationer, andningssymtom och oförutsägbara farmakologiska effekter [B306].
Vanliga förfalskningar
- Syntetiska tillsatser: Syntetisk linalool eller linalylacetat i “lavendel”-olja [B307]
- Spädningsmedel Propylenglykol, Dipro-pylenglykolmetyleter (DPM) [B308]
- Billigare oljor: Lavandin (Lavandula x intermedia) säljs som äkta lavendel (Lavandula angustifolia) [B309]
- Bekämpningsmedel Restmängder av bekämpningsmedel, ogräsmedel i icke-ekologiska oljor [B310]
Säkerhetsrekommendation
Använd endast GC/MS-certifierade, 100 % rena, ekologiska eteriska oljor från pålitliga leverantörer. Begär GC/MS-certifikat för varje parti [B311].
diskussion
Evidensläge
Bevisen för eteriska oljor vid ASD är heterogena och av övervägande låg kvalitet. Den kliniska litteraturen inkluderar en negativ randomiserad kontrollerad studie (bergamott vid akut ångest) [B200], små pilotstudier med metodologiska begränsningar (lavendel vid ABA-efterlevnad) [B198], observationsstudier (aromaterapimassage) [B199] och fallrapporter [B312]. Ingen studie har demonstrerat effekt för ASD:s kärnsymtom (social kommunikation, repetitivt beteende) med tillräcklig evidensgrad [B313].
I kontrast är den prekliniska evidensen robust: Flera studier visar att specifika terpener (linalool, β-karyofyllen, boswellinsyror, 1,8-cineol) angriper ASD-relevanta molekylära mål (GABA-A-receptorer, CB2-receptorer, Nrf2-vägen, neuroinflammation) och förbättrar ASD-liknande beteenden i djurmodeller. [B152], [B161], [B174], [B183], [B197]. Denna diskrepans mellan preklinisk trovärdighet och klinisk evidens är typisk för komplementära terapier och återspeglar brist på forskningsfinansiering, metodologiska utmaningar och translationsluckor. [B314].
Mekanistisk trovärdighet
Den mekanistiska trovärdigheten för eteriska oljor vid ASD finns, baserat på konvergens mellan ASD-patofysiologi och terpeners verkningsmekanismer:
GABA/Glutamat E/I-obalans
Linalool och 1,8-cineol modulerar GABA-A-receptorer vid bensodiazepinbindningsstället, vilket kan förstärka hämmande neurotransmission och normalisera E/I-balansen. [B152], [B183]. Dessa mekanismer är analoga med bumetanid, som har visat måttliga förbättringar vid ASD i kliniska studier [B52].
Neuroinflammation
Beta-karyofyllen (CB2-agonist) och boswelliasyror (Nrf2-aktivator) minskar mikrogliaaktivering, proinflammatoriska cytokiner och oxidativ stress i prekliniska modeller [B161], [B174]. Eftersom neuroinflammation är en etablerad patofysiologisk mekanism vid ASD [B77], skulle dessa terpener kunna vara användbara för patienter med neuroinflammation (förhöjda perifera cytokiner, PET-undersökning som visar mikrogliaaktivering) [B315].
Serotonin/Dopamin
Linalool och Ylang-Ylang modulerar serotonerga och dopaminerga system i prekliniska studier [B154], [B197]. Dessa mekanismer skulle kunna förbättra komorbida symtom (ångest, humör), även om effekter på kärnsymtom är osannolika [B316].
Oxytocin
Linalool ökar den intracellulära kalciumkoncentrationen i oxytocinneuroner [B155]. Eftersom oxytocin reglerar GABAerg utveckling och förbättrade ASD-liknande fenotyper i prekliniska studier [B50], skulle linalool kunna modulera oxytongerga signalvägar indirekt [B317].
Tarm-hjärna-axeln
Trots avsaknad av direkta bevis, kan antiinflammatoriska terpener (β-karyofyllen, boswelliasyror) minska systemisk inflammation och indirekt modulera tarm-hjärna-axelns signalvägar [B318]. Denne hypotese krever eksperimentell validering [B319].
Begränsningar
Små studier
De flesta ASD-specifika studier har provstorlekar på <20, vilket ger otillräcklig styrka för att upptäcka måttliga effekter [B320]. Meta-analyser är inte möjliga på grund av heterogenitet [B321].
Inga randomiserade kontrollerade studier för kärnreaktioner
Det finns inga randomiserade kontrollerade studier som undersöker effekten av eteriska oljor för kärnsymptom vid ASD (social kommunikation, repetitiva beteenden). [B322]. Den enda identifierade RCT (bergamott) var negativ för akut ångest [B200].
Metodologisk heterogenitet
Studierna varierar i oljeval, dosering, administrationssätt (inhalation, massage, topisk), exponeringstid, utfallsmått och populationer [B323]. Denna heterogenitet förhindrar syntes och jämförbarhet [B324].
Saknade biomarkörer
Ingen studie stratifierar patienter efter neurobiologiska subtyper (t.ex. neuroinflammatoriska signaturer, E/I-obalansmarkörer) eller använder objektiva biomarkörer som utfall. [B325]. Precisionsmedicinska metoder kan identifiera svarare [B326].
Publikationsbias
Positiva fallrapporter och små pilotstudier publiceras oftare än negativa studier, vilket kan leda till en överskattning av effekter [B327].
Översättningslucka
Prekliniska modeller (VPA-råttor, MIA-möss) replikerar inte den fulla komplexiteten hos mänsklig ASD-heterogenitet [B328]. Terpendoser i djurstudier är ofta högre än vad som kan uppnås vid human inhalationsexponering [B329].
Placeboeffekter
Aromaterapi är förknippad med starka förväntningseffekter; blindning är svår på grund av påtagliga lukter [B330]. Bergamott-RCT använde vatten som placebo, vilket innebär otillräcklig blindning [B200].
Säkerhetsdata Långtidsdata om säkerheten vid kronisk inhalationsexponering hos barn saknas [B331]. De flesta säkerhetsdata kommer från topisk eller oral applicering [B332].
Utsikt
Forskningsprioriteringar:
- Högkvalitativa RCT:er: Randomiserade, dubbelblinda, placebokontrollerade studier med tillräcklig styrka (n > 100), standardiserade oljepreparat (GC/MS-certifierade), objektiva utfallsmått (biomarkörer, aktigrafi, eye-tracking) och längre interventionsperioder (8–12 veckor) [B333].
- Stratifiering efter subtyper: Identifiering av responders baserat på neurobiologiska subtyper (neuroinflammationsmarkörer, E/I-obalans, mikrobiomprofiler) [B334]. Precisionsmedicinska angreppssätt kan öka effektstorlekar [B335].
- Mekanistiska studier: Humana studier med biomarkörutfall (cytokinprofiler, neuroavbildning, EEG, mikrobiomanalyser) före och efter intervention, för att validera mekanismer [B336].
- Farmakokinetik Humana farmakokinetiska studier för inhalationsexponering för terpener: plasmakoncentrationer, penetration till centrala nervsystemet, metabolism, dos-respons-relationer [B337].
- Kombinationsterapin: Undersökning av synergier mellan eteriska oljor och etablerade interventioner (beteendeterapi, farmakoterapi, mikrobiominterventioner) [B338].
- Långsiktig säkerhet: Prospektiv kohortstudie om långtidssäkerheten vid kronisk inhalations exponering hos barn [B339].
- Personlig aromaterapi: Utveckling av algoritmer för oljeval baserat på individuella symtomprofiler, preferenser och neurobiologiska markörer [B340].
Kliniska implikationer
Baserat på den nuvarande evidensen kan eteriska oljor övervägas som en del av en multimodal behandlingsansats för komorbida symtom (ångest, sömnstörningar, sensorisk dysreglering) vid ASD, men inte som monoterapi eller ersättning för evidensbaserade kärninterventioner. [B341]. Kliniker bör:
- Kommunicera realistiska förväntningar: Eteriska oljor kan lindra komorbida symptom, men inte bota kärnsymptom [B342]
- Ta hänsyn till individuella toleranser och preferenser [B343]
- Rekommendera högkvalitativa, GC/MS-certifierade oljor [B344]
- Ta hänsyn till säkerhetsaspekter (epilepsi, ålder, läkemedelsinteraktioner) [B345]
- Effektövervakning och behandlingsjustering [B346]
Slutsats
Eteriska oljor utgör ett mekanistiskt plausibelt men kliniskt otillräckligt validerat adjunktivt terapeutiskt alternativ för autismspektrumtillstånd. Den neurobiologiska heterogeniteten hos ASD – kännetecknad av GABA/glutamat E/I-obalans, neuroinflammation, oxidativ stress, förändringar i tarm-hjärna-axeln och synaptisk dysfunktion – erbjuder flera terapeutiska mål som specifika terpener kan angripa.
Prekliniska bevis visar att linalool (GABA-A-modulering, serotonerg reglering), β-karyofyllen (CB2-medierad minskning av neuroinflammation), boswelliasyror (aktivering av Nrf2/HO-1), 1,8-cineol (ångestdämpande GABAerga effekter) och ylang-ylang (serotonin/dopamin-modulering) modulerar ASD-relaterade patofysiologiska mekanismer och förbättrar ASD-liknande beteenden i djurmodeller. Denna mekanistiska trovärdighet stöds av traditionella användningsområden och begränsade humana data för ångest, sömn och humör.
De kliniska beläggen vid AST är dock av låg kvalitet: en negativ RCT (bergamott), små pilotstudier med metodologiska begränsningar (lavendel vid ABA-compliance) och observationsstudier. Ingen studie påvisar effekt för AST:s kärnsymtom med tillräcklig evidenskvalitet. Högkvalitativa, randomiserade kontrollerade studier med standardiserade preparat, objektiva utfall och tillräcklig styrka saknas.
För Organik Aromas Nebulizer 3.0 utvecklas doseringsrekommendationer specifika för rumsstorlek (2–10 droppar för 10–60 m², 15–30 minuters diffusion, 30–60 minuters pauser), med tidpunktsanpassade protokoll (aktiverande morgon, lugnande kväll) och ASD-specifika hänsyn (gradvis introduktion, preferenstester, sensorisk överkänslighet). Säkerhetsaspekter inkluderar kontraindikationer vid epilepsi (kampherhaltiga oljor), barn <3 år (eukalyptus), potentiella interaktioner med antipsykotika (bergaptenhaltiga oljor) och behovet av GC/MS-certifierade, rena oljor.
Eteriska oljor kan övervägas som en del av ett multimodalt terapeutiskt tillvägagångssätt för komorbida symtom (ångest, sömnstörningar, sensorisk dysreglering) vid ASD, men de ersätter inte evidensbaserade kärninterventioner (beteendeterapi, strukturerad utbildning). Framtida forskning bör prioritera högkvalitativa RCT, stratifiering baserad på neurobiologiska subtyper, mekanistiska biomarkörstudier och långsiktiga säkerhetsdata för att validera den terapeutiska potentialen hos eteriska oljor vid ASD och för att utveckla precisionsmedicinska tillvägagångssätt.
Gemensam slutsats
Övergripande insikter: ADHD och autism
Båda neurodevelopmentala störningarna, ADHD och autismspektrumstörning (ASD), delar grundläggande neurobiologiska likheter som gör en adjuvant terapi med eteriska oljor trovärdig för båda indikationerna.
Gemensamma neurobiologiska målstukturrer:

Gemensam rekommendation för användning (Organik Aromas Nebulizer 3.0):
För båda indikationerna gäller Venturi-kallförsto principer utan vatten eller värme som optimal appliceringsform: – Ingen termisk nedbrytning av terpenerna – Exakt partikelstorlek för olfaktorisk absorption (< 5 µm) – GC/MS-rena oljor bibehåller sin molekylära integritet
Viktig meddelande
Adjuvant behandling med eteriska oljor ersätter inte evidensbaserad standardbehandling (metylfenidat vid ADHD, ABA/beteendeterapi vid ASD). Den utgör ett komplementärt tillägg som bör anpassas individuellt och genomföras under professionell handledning. Evidensläget är fortfarande begränsat för båda indikationerna; större RCT:er behövs.
Referens ADHD
Chen et al. (2008). Effekten av kanel-aromaterapi för barn med ADHD.https://pmc.ncbi.nlm.nih.gov/articles/PMC8538900/
[A2] Chen et al. Effekten av att applicera kanel-aromaterapi för barn med uppmärksamhetsunderskott hyperaktivitetsstörning.https://pmc.ncbi.nlm.nih.gov/articles/PMC8538900/
[A3] Godfrey (2001). Eteriska oljors roll i behandling och hantering av uppmärksamhetsstörning med hyperaktivitet. International Journal of Aromatherapy.https://doi.org/10.1016/S0962-4562(01 )80035-7
[A4] Godfrey. Etiska oljor: Komplementär behandling för ADHD (neuropsykiatrisk funktionsnedsättning).https://www.researchgate.net/publication/248871372_The_role_of_essential_oils_in_the_treatment_and_management_of_attention_deficit_hyperactive_disorder
[A5] Godfrey. ESSENTIELLA OLJORS ROLL VID BEHANDLING OCH HANTERING AV UPMÄRKSAMHETSSTÖRNING MED HYPERAKTIVITET (ADHD).https://www.researchgate.net/publication/248871372_The_role_of_essential_oils_in_the_treatment_and_management_of_attention_deficit_hyperactive_disorder
[A6] Ding et al. (1994). Farmakokinetik och in vivo-specificitet för [LLC]dl-treo-metylfenidat för det presynaptiska dopaminergiska neuronet. Synapse.https://doi.org/10.1002/SYN.890180207
[A7] Volkow et al. (2002). Methylfenidats verkningsmekanism: insikter från PET-avbildningsstudier. Journal of Attention Disorders.https://doi.org/10.1177/070674370200601S05
[A8] Silva (2024). Aromaterapi vid vård av atypiskt barn med ADHD och atopiskt eksem.https://doi.org/10.62435/2965-7253.bjhae.2024.iisinaroma1
[A9] Volkow et al. (1998). Dopamintransportörsbeläggningar i den mänskliga hjärnan orsakade av terapeutiska doser av oralt metylfenidat. American Journal of Psychiatry.https://doi.org/10.1176/AJP.155.10.1325
[A10] Jacob et al. (2024). Aromaterapiens användning inom studenthälsovården.https://doi.org/10.37885/240717094
[A11] Dipasquale et al. (2020). Att reda ut effekterna av metylfenidat på de dopaminerga och noradrenerga funktionella kretsarna. bioRxiv.https://doi.org/10.1101/2020.03.09.983528
[A12] Volkow et al. (2002). Dopaminets roll i metylfenidatets terapeutiska och förstärkande effekter hos människor: resultat från bildstudiier. European Neuropsychopharmacology.https://doi.org/10.1016/S0924-977X(02 )00104-9
[A13] Spencer et al. (2006). PET-studie som undersöker farmakokinetik, upptäckt och gillande samt dopamintransportörreceptorens upptagningsgrad av kort- och långverkande oralt metylfenidat. American Journal of Psychiatry.https://doi.org/10.1176/APPI.AJP.163.3.387
[A14] Castillo et al. (2018). Eteriska oljor som modifierare av mänskligt beteende. Tropiska och subtropiska agroekosystem.https://www.revista.ccba.uady.mx/ojs/index.php/TSA/article/view/2583
[A15] Crunelle et al. (2014). Dopamintransportörernas ockupans av metylfenidat och impulsivitet vid ADHD hos vuxna. British Journal of Psychiatry.https://doi.org/10.1192/BJP.BP.113.132977
[A16] Ding et al. (1997). Kirala läkemedel: jämförelse av farmakokinetiken för [11C]d-treo och l-treo-metylfenidat i mänsklig och babianhjärna. Psychopharmacology.https://doi.org/10.1007/S002130050267
[A17] Spencer et al. (2009). En PET-studie som undersöker farmakokinetik och dopamintransportörbeläggning av två långverkande formuleringar av metylfenidat hos vuxna. International Journal of Molecular Medicine.https://doi.org/10.3892/IJMM_00000339
[A18] Marcel m.fl. (2024). Striatal dopaminsystems responsivitet på metylfenidat – en inomindividuell I-123-β-CIT-SPECT-studie hos pojkar med adhd.https://doi.org/10.5283/epub.57629
[A19] Kennedy et al. (2018). Flyktiga terpener och hjärnfunktion: Undersökning av de kognitiva effekterna och humörseffekterna av Mentha × Piperita L. eterisk olja med in vitro-egenskaper relevanta för centrala nervsystemets funktion. Nutrients.https://doi.org/10.3390/NU10081029
[A20] Batiha et al. (2023). En översikt av de bioaktiva komponenterna och farmakologiska egenskaperna hos Lavandula-arter. Naunyn-schmiedebergs Archives of Pharmacology.https://doi.org/10.1007/s00210-023-02392-x
[A21] Guzmán-Gutiérrez et al. (2015). Linanol och β-pinen utövar sin antidepressiva aktivitet genom monoaminerg väg.https://pubmed.ncbi.nlm.nih.gov/25771248/
[A22] Martinez et al. (2020). Ventrala striatum stöder metylfenidatets terapeutiska effekter på impulsiva val som uttrycks i en uppgift om tidspresent. Scientific reports.https://doi.org/10.1038/s41598-020-57595-6
[A23] López et al. (2017). Utforskning av Farmakologiska Mekanismer hos Lavendel (Lavandula angustifolia) Eterisk Olja på Centrala Nervsystemets Mål. Frontiers in Pharmacology.https://doi.org/10.3389/FPHAR.2017.00280
[A24] Rathnayake (2025). “Neuroaromater”: Aromaterapi och det limbiska stressresponset.https://doi.org/10.4038/jssnisd.v3i1.28
[A25] Aster et al. (2021). Striatala dopaminsystemets responsivitet på metylfenidat – en inomindividuell I-123-β-CIT-SPECT-studie hos barn och ungdomar med ADHD. medRxiv.https://doi.org/10.1101/2021.11.19.21265993
[A26] Cardoso et al. (2025). Lavendeloljans inverkan på det mänskliga nervsystemet: En översikt. Deleted Journal.https://doi.org/10.62435/2965-7253.bjhae.2025.bjhae22
[A27] Moss et al. (2003). Aromas av rosmarin- och lavendel eteriska oljor påverkar kognition och humör selektivt hos friska vuxna. International Journal of Neuroscience.https://doi.org/10.1080/00207450390161903
[A28] Sinha et al. (2005). Komplementär och alternativ medicin vid behandling av barn med ADHD. Journal of Paediatrics and Child Health.https://doi.org/10.1111/J.1440-1754.2005.00530.X
[A29] Springer et al. (2018). ADHD: Livslängd och effekter från ungdomsåren genom vuxenlivet.https://dergipark.org.tr/en/pub/pgy/548052.xml
[A30] Colzato et al. (2014). Attentional control in the attentional blink is modulated by odor. Attention Perception & Psychophysics.https://doi.org/10.3758/S13414-014-0733-0
A31. Kennedy, D. O., Scholey, A., Tildesley, N. T. J., Perry, E. K., & Wesnes, K. A. (2002). Modulation av humör och kognitiv prestation efter akut administrering av Melissa officinalis (citronmeliss). Farmakologi, biokemi och beteende, 72(4), 953–964. https://doi.org/10.1016/S0091-3057(02)00777-3
A32. Müller, S. F., & Klement, S. (2006). En kombination av valeriana och citronmeliss är effektiv vid behandling av rastlöshet och insomni hos barn. Fytomedicin, 13(6), 383–387. https://doi.org/10.1016/j.phymed.2006.01.013
A33. Scholey, A., Gibbs, A., Neale, C., et al. (2014). Antistresseffekter av livsmedel innehållande citronmeliss. Näringsämnen, 6(11), 4805–4821. https://doi.org/10.3390/nu6114805
A34. Översikt av neuroprotektiva effekter av eteriska oljor inklusive Eucalyptus globulus och Jasminum sambac på kognitiv funktion och Alzheimerrelaterad patologi. (2022). Neuroprotektiva eteriska oljor Översikt.
Referens ASD
[B1] Maenner MJ, Shaw KA, Bakian AV, et al. Förekomst och karakteristika för autism i spektrum bland barn 8 år — Autism and Developmental Disabilities Monitoring Network, 11 platser, USA, 2018. MMWR Surveill Summ. 2021;70(11):1-16.https://doi.org/10.15585/mmwr.ss7011a1
[B2] Zeidan J, Fombonne E, Scorah J, et al. Global prevalence of autism: A systematic review update. Autism Res. 2022;15(5):778-790.https://doi.org/10.1002/aur.2696
[B3] Lord C, Elsabbagh M, Baird G, Veenstra-Vanderweele J. Autism spectrum disorder. Lancet. 2018;392(10146):508-520.https://doi.org/10.1016/S0140-6736(18 )31129-2
[B4] Masi A, DeMayo MM, Glozier N, Guastella AJ. En översikt över autismdiagnosen, heterogenitet och behandlingsalternativ. Neurosci Bull. 2017;33(2):183-193.https://doi.org/10.1007/s12264-017-0100-y
[B5] American Psychiatric Association. Diagnostic and Statistical Manual of Mental Disorders, Fifth Edition (DSM-5). Arlington, VA: American Psychiatric Publishing; 2013.https://www.psychiatry.org/psychiatrists/practice/dsm
[B6] Marco EJ, Hinkley LB, Hill SS, Nagarajan SS. Sensorisk bearbetning vid autism: en översikt av neurofysiologiska fynd. Pediatr Res. 2011;69(5 Pt 2):48R-54R.https://doi.org/10.1203/PDR.0b013e3182130c54
[B7] Baum SH, Stevenson RA, Wallace MT. Beteendemässiga, perceptuella och neurala förändringar i sensorisk och multisensorisk funktion vid autism. Prog Neurobiol. 2015;134:140-160.https://doi.org/10.1016/j.pneurobio.2015.09.007
[B8] van Steensel FJ, Bögels SM, Perrin S. Ångeststörningar hos barn och unga med autismspektrumstörningar: en metaanalys. Clin Child Fam Psychol Rev. 2011;14(3):302-317.https://doi.org/10.1007/s10567-011-0097-0
[B9] Souders MC, Mason TB, Valladares O, et al. Beteenden kring sömn och sömnkvalitet hos barn med autismspektrumtillstånd. Sleep. 2009;32(12):1566-1578.https://doi.org/10.1093/sleep/32.12.1566
[B10] McElhanon BO, McCracken C, Karpen S, Sharp WG. Gastrointestinal symptoms in autism spectrum disorder: a meta-analysis. Pediatrics. 2014;133(5):872-883.https://doi.org/10.1542/peds.2013-3995
[B11] Geschwind DH, State MW. Genjakt vid autism-spektrumstörning: på vägen mot precisionsmedicin. Lancet Neurol. 2015;14(11):1109-1120.https://doi.org/10.1016/S1474-4422(15 )00044-7
[B12] De Rubeis S, He X, Goldberg AP, et al. Synaptiska, transkriptionella och kromatingener som störs vid autism. Nature. 2014;515(7526):209-215.https://doi.org/10.1038/nature13772
[B13] Sandin S, Lichtenstein P, Kuja-Halkola R, et al. Ärftligheten för autism-spektrumstörning. JAMA. 2017;318(12):1182-1184.https://doi.org/10.1001/jama.2017.12141
[B14] Rubenstein JL, Merzenich MM. Modell av autism: ökat förhållande mellan excitation/hämning i viktiga neurala system. Genes Brain Behav. 2003;2(5):255-267.https://doi.org/10.1034/j.1601-183x.2003.00037.x
[B15] Vargas DL, Nascimbene C, Krishnan C, Zimmerman AW, Pardo CA. Neuroglial aktivering och neuroinflammation i hjärnan hos patienter med autism. Ann Neurol. 2005;57(1):67-81.https://doi.org/10.1002/ana.20315
[B16] Rossignol DA, Frye RE. Bevis som kopplar oxidativ stress, mitokondriell dysfunktion och inflammation i hjärnan hos personer med autism. Front Physiol. 2014;5:150.https://doi.org/10.3389/fphys.2014.00150
[B17] Nelson SB, Valakh V. Excitatorisk/inhibitorisk balans och krets homeostas vid autismspektrumstörningar. Neuron. 2015;87(4):684-698.https://doi.org/10.1016/j.neuron.2015.07.033
[B18] Lee E, Lee J, Kim E. Excitation/Inhibition Imbalance in Animal Models of Autism Spectrum Disorders. Biol Psychiatry. 2017;81(10):838-847.https://doi.org/10.1016/j.biopsych.2016.05.011
[B19] Horder J, Lavender T, Mendez MA, et al. Minskat subkortikalt glutamat/glutamin hos vuxna med autism-spektrumstörningar: en [¹H]MRS-studie. Transl Psychiatry. 2013;3:e279.https://doi.org/10.1038/tp.2013.53
[B20] Gaetz W, Bloy L, Wang DJ, et al. GABA-uppskattning i hjärnor hos barn med autism som spektrum: mätprecision och regional variation i barken. Neuroimage. 2014;86:1-9.https://doi.org/10.1016/j.neuroimage.2013.05.068
[B21] Rojas DC, Singel D, Steinmetz S, Hepburn S, Brown MS. Minskad vänster perisylvisk GABA-koncentration hos barn med autism och opåverkade syskon. Neuroimage. 2014;86:28-34.https://doi.org/10.1016/j.neuroimage.2013.01.045
[B22] Amaral DG, Schumann CM, Nordahl CW. Neuroanatomy of autism. Trends Neurosci. 2008;31(3):137-145.https://doi.org/10.1016/j.tins.2007.12.005
[B23] Courchesne E, Pierce K. Why the frontal cortex in autism might be talking only to itself: local over-connectivity but long-distance disconnection. Curr Opin Neurobiol. 2005;15(2):225-230.https://doi.org/10.1016/j.conb.2005.03.001
[B24] Baron-Cohen S, Ring HA, Bullmore ET, Wheelwright S, Ashwin C, Williams SC. Amygdalateorin om autism. Neurosci Biobehav Rev. 2000;24(3):355-364.https://doi.org/10.1016/s0149-7634(00 )00011-7
[B25] Estes ML, McAllister AK. Maternal immune activation: Implications for neuropsychiatric disorders. Science. 2016;353(6301):772-777.https://doi.org/10.1126/science.aag3194
[B26] Matta SM, Hill-Yardin EL, Crack PJ. Neuroinflammationens inverkan på autismspektrumstörning. Brain Behav Immun. 2019;79:75-90.https://doi.org/10.1016/j.bbi.2019.04.037
[B27] Careaga M, Murai T, Bauman MD. Maternal Immune Activation and Autism Spectrum Disorder: From Rodents to Nonhuman and Human Primates. Biol Psychiatry. 2017;81(5):391-401.https://doi.org/10.1016/j.biopsych.2016.10.020
Chauhan A, Chauhan V. Oxidative stress in autism. Pathophysiology. 2006;13(3):171-181.https://doi.org/10.1016/j.pathophys.2006.05.007
[B29] Rose S, Melnyk S, Pavliv O, et al. Evidence of oxidative damage and inflammation associated with low glutathione redox status in the autism brain. Transl Psychiatry. 2012;2:e134.https://doi.org/10.1038/tp.2012.61
[B30] Kang DW, Adams JB, Gregory AC, et al. Microbiota Transfer Therapy förändrar tarmekosystemet och förbättrar gastrointestinala och autism symptom: en öppen studie. Microbiome. 2017;5(1):10.https://doi.org/10.1186/s40168-016-0225-7
[B31] Vuong HE, Hsiao EY. Tarmflorans nya roller vid autism. Biol Psychiatry. 2017;81(5):411-423.https://doi.org/10.1016/j.biopsych.2016.08.024
[B32] Sharon G, Cruz NJ, Kang DW, et al. Human Gut Microbiota from Autism Spectrum Disorder Promote Behavioral Symptoms in Mice. Cell. 2019;177(6):1600-1618.e17.https://doi.org/10.1016/j.cell.2019.05.004
[B33] Sgritta M, Dooling SW, Buffington SA, et al. Mekanismer som ligger bakom mikrobiellt medierade förändringar i socialt beteende i musmodeller av autismspektrumstörning. Neuron. 2019;101(2):246-259.e6.https://doi.org/10.1016/j.neuron.2018.11.018
[B34] Reichow B, Hume K, Barton EE, Boyd BA. Tidig intensiv beteendeintervention (EIBI) för små barn med autismspektrumstörningar (ASD). Cochrane Database Syst Rev. 2018;5(5):CD009260.https://doi.org/10.1002/14651858.CD009260.pub3
[B35] Dawson G, Rogers S, Munson J, et al. Randomiserad, kontrollerad studie av en intervention för småbarn med autism: Early Start Denver-modellen. Pediatrics. 2010;125(1):e17-23.https://doi.org/10.1542/peds.2009-0958
[B36] McCracken JT, McGough J, Shah B, et al. Risperidon hos barn med autism och allvarliga beteendeproblem. N Engl J Med. 2002;347(5):314-321.https://doi.org/10.1056/NEJMoa013171
[B37] Marcus RN, Owen R, Kamen L, m.fl. En placebokontrollerad studie med fast dos av aripiprazol hos barn och ungdomar med irritabilitet associerad med autism. J Am Acad Child Adolesc Psychiatry. 2009;48(11):1110-1119.https://doi.org/10.1097/CHI.0b013e3181b76658
[B38] Scahill L, McDougle CJ, Aman MG, et al. Effekter av risperidon och föräldraträning på adaptiv funktion hos barn med genomgripande utvecklingsstörningar och allvarliga beteendeproblem. J Am Acad Child Adolesc Psychiatry. 2012;51(2):136-146.https://doi.org/10.1016/j.jaac.2011.11.010
[B39] Rossignol DA, Frye RE. Melatonin vid autism-spektrumstörningar: en systematisk översikt och metaanalys. Dev Med Child Neurol. 2011;53(9):783-792.https://doi.org/10.1111/j.1469-8749.2011.03980.x
[B40] Hollander E, Soorya L, Chaplin W, et al. En dubbelblind placebokontrollerad studie av fluoxetin för repetitiva beteenden och global svårighetsgrad vid autism-spektrumstörningar hos vuxna. Am J Psychiatry. 2012;169(3):292-299.https://doi.org/10.1176/appi.ajp.2011.10050764
[B41] Spencer D, Marshall J, Post B, m.fl. Psykofarmakabehandling och kombinationsbehandling hos barn med autism. Pediatrics. 2013;132(5):833-840.https://doi.org/10.1542/peds.2012-3774
[B42] Koulivand PH, Khaleghi Ghadiri M, Gorji A. Lavendel och nervsystemet. Evid Based Complement Alternat Med. 2013;2013:681304.https://doi.org/10.1155/2013/681304
[B43] Sowndhararajan K, Kim S. Influence of Fragrances on Human Psychophysiological Activity: With Special Reference to Human Electroencephalographic Response. Sci Pharm. 2016;84(4):724-751.https://doi.org/10.3390/scipharm84040724
[B44] Ali B, Al-Wabel NA, Shams S, Ahamad A, Khan SA, Anwar F. Eteriska oljor som används inom aromaterapi: En systematisk översikt. Asian Pac J Trop Biomed. 2015;5(8):601-611.https://doi.org/10.1016/j.apjtb.2015.05.007
[B45] Sohal VS, Rubenstein JLR. Excitation-inhibition balance som en ram för att undersöka mekanismer vid neuropsykiatriska sjukdomar. Mol Psychiatry. 2019;24(9):1248-1257.https://doi.org/10.1038/s41380-019-0426-0
[B46] Antoine MW, Langberg T, Schnepel P, Feldman DE. Ökat förhållande mellan excitation och inhibition stabiliserar synapser och krets-excitabilitet i fyra mustarter av autism. Neuron. 2019;101(4):648-661.e4.https://doi.org/10.1016/j.neuron.2018.12.026
[B47] Fatemi SH, Halt AR, Stary JM, Kanodia R, Schulz SC, Realmuto GR. Glirmsyraddekarboxylas 65 och 67 kDa-proteiner är reducerade i autistiska parietala och cerebellära kortex. Biol Psychiatry. 2002;52(8):805-810.https://doi.org/10.1016/s0006-3223(02 )01430-0
[B48] Yip J, Soghomonian JJ, Blatt GJ. Minskade GAD67 mRNA-nivåer i cerebellära Purkinjeceller vid autism: patofysiologiska implikationer. Acta Neuropathol. 2007;113(5):559-568.https://doi.org/10.1007/s00401-006-0176-3
[B49] Ben-Ari Y. Excitatoriska GABA-effekter under utveckling: naturens nytta. Nat Rev Neurosci. 2002;3(9):728-739.https://doi.org/10.1038/nrn920
[B50] Tyzio R, Nardou R, Ferrari DC, et al. Oxytocin-medierad GABA-hämning under förlossningen dämpar autismgenesen hos gnäggavkommor. Science. 2014;343(6171):675-679.https://doi.org/10.1126/science.1247190
[B51] Lemonnier E, Degrez C, Phelep M, et al. En randomiserad kontrollerad studie av bumetanid vid behandling av autism hos barn. Transl Psychiatry. 2012;2:e202.https://doi.org/10.1038/tp.2012.124
[B52] Lemonnier E, Villeneuve N, Sonie S, et al. Effekter av bumetanid på neurobeteendefunktionen hos barn och ungdomar med autismspektrumstörningar. Transl Psychiatry. 2017;7(3):e1056.https://doi.org/10.1038/tp.2017.10
[B53] Carlson GC. Glutamatreceptor dysfunktion och läkemedelsmål över modeller av autism. Pharmacol Biochem Behav. 2012;100(4):850-854.https://doi.org/10.1016/j.pbb.2011.02.003
[B54] Uzunova G, Pallanti S, Hollander E. Excitatorisk/inhibitorisk obalans vid autismspektrumstörningar: Implikationer för interventioner och terapier. World J Biol Psychiatry. 2016;17(3):174-186.https://doi.org/10.3109/15622975.2015.1085597
[B55] Yoo HJ, Cho IH, Park M, Yang SY, Kim SA. Familjebaserad association av GRIN2A och GRIN2B med koreanska autismspektrumstörningar. Neurosci Lett. 2012;512(2):89-93.https://doi.org/10.1016/j.neulet.2012.01.061
[B56] Silverman JL, Tolu SS, Barkan CL, Crawley JN. Repetitivt självputsande beteende hos BTBR-musmodellen för autism blockeras av mGluR5-antagonisten MPEP. Neuropsychopharmacology. 2010;35(4):976-989.https://doi.org/10.1038/npp.2009.201
[B57] Muller CL, Anacker AMJ, Veenstra-VanderWeele J. Serotoninsystemet vid autism:[1]spektrumstörning: från biomarkör till djurmodeller. Neuroscience. 2016;321:24-41.https://doi.org/10.1016/j.neuroscience.2015.11.010
[B58] Pavăl D. En dopaminhypotes för autismspektrumstörning. Dev Neurosci. 2017;39(5):355-360.https://doi.org/10.1159/000478725
[B59] Schain RJ, Freedman DX. Studier av 5-hydroxiindolmetabolism hos autistiska och andra mentalt retarderade barn. J Pediatr. 1961;58:315–320.https://doi.org/10.1016/s0022-3476(61 )80261-8
[B60] Cook EH Jr, Leventhal BL. Serotoninsystemet vid autism. Curr Opin Pediatr. 1996;8(4):348-354.https://doi.org/10.1097/00008480-199608000-00008
[B61] Chugani DC, Muzik O, Behen M, et al. Utvecklingsförändringar i hjärnans serotoninsyntesförmåga hos autistiska och icke-autistiska barn. Ann Neurol. 1999;45(3):287-295.https://doi.org/10.1002/1531-8249(199903 )45:33.0.co;2-9
[B62] Devlin B, Cook EH Jr, Coon H, et al. Autism och serotonintransportören: det långa och korta av det. Mol Psykiatri. 2005;10(12):1110-1116.https://doi.org/10.1038/sj.mp.4001724
[B63] Whitaker-Azmitia PM. Beteendemässiga och cellulära konsekvenser av ökad serotonerg aktivitet under hjärnans utveckling: en roll i autism? Int J Dev Neurosci. 2005;23(1):75-83.https://doi.org/10.1016/j.ijdevneu.2004.07.022
King BH, Hollander E, Sikich L, et al. Bristande effekt av citalopram hos barn med autismspektrumstörningar och höga nivåer av repetitivt beteende: citalopram ineffektivt hos barn med autism. Arch Gen Psychiatry. 2009;66(6):583-590.https://doi.org/10.1001/archgenpsychiatry.2009.30
[B65] Dichter GS, Damiano CA, Allen JA. Dysfunktion i belöningskretsloppet vid psykiatriska och neuropsykiatriska störningar och genetiska syndrom: djurmodeller och kliniska fynd. J Neurodev Disord. 2012;4(1):19.https://doi.org/10.1186/1866-1955-4-19
[B66] Kohls G, Schulte-Rüther M, Nehrkorn B, et al. Belöningssystemets dysfunktion vid autism. Soc Cogn Affect Neurosci. 2013;8(5):565-572.https://doi.org/10.1093/scan/nss033
[B67] Delmonte S, Balsters JH, McGrath J, et al. Social and monetary reward processing in autism spectrum disorders. Mol Autism. 2012;3(1):7.https://doi.org/10.1186/2040-2392-3-7
[B68] Nakamura K, Sekine Y, Ouchi Y, et al. Serotonin- och dopamintransportörbindningar i hjärnan hos vuxna med högfungerande autism. Arch Gen Psychiatry. 2010;67(1):59-68.https://doi.org/10.1001/archgenpsychiatry.2009.137
[B69] Staal WG. Autism, DRD3 och repetitivt och stereotypiskt beteende, en översikt över nuvarande kunskap. Eur Neuropsychopharmacol. 2015;25(9):1421-1426.https://doi.org/10.1016/j.euroneuro.2014.08.011
[B70] Gunaydin LA, Grosenick L, Finkelstein JC, m.fl. Naturliga neurala projektionsdynamik som ligger till grund för socialt beteende. Cell. 2014;157(7):1535-1551.https://doi.org/10.1016/j.cell.2014.05.017
[B71] Insel TR. Utmaningen med översättning inom social neurovetenskap: en översikt av oxytocin, vasopressin och tillgivenhetsbeteende. Neuron. 2010;65(6):768-779.https://doi.org/10.1016/j.neuron.2010.03.005
[B72] Modahl C, Green L, Fein D, et al. Plasmatoxytocinivåer hos autistiska barn. Biol Psychiatry. 1998;43(4):270-277.https://doi.org/10.1016/s0006-3223(97 )00439-3
[B73] Gregory SG, Connelly JJ, Towers AJ, et al. Genomiska och epigenetiska bevis för oxytocinreceptordefekt vid autism. BMC Med. 2009;7:62.https://doi.org/10.1186/1741-7015-7-62
[B74] Guastella AJ, Einfeld SL, Gray KM, et al. Intranasal oxytocin förbättrar igenkänning av känslor hos ungdomar med autismspektrumtillstånd. Biol Psychiatry. 2010;67(7):692-694.https://doi.org/10.1016/j.biopsych.2009.09.020
[B75] Parker KJ, Oztan O, Libove RA, et al. Intranasal oxytocinbehandling för sociala brister och biomarkörer för respons hos barn med autism. Proc Natl Acad Sci U S A. 2017;114(30):8119-8124.https://doi.org/10.1073/pnas.1705521114
[B76] Peñagarikano O, Lázaro MT, Lu XH, et al. Exogent och framkallat oxytocin återställer socialt beteende i Cntnap2-musmodellen av autism. Sci Transl Med. 2015;7(271):271ra8.https://doi.org/10.1126/scitranslmed.3010257
[B77] Pardo CA, Vargas DL, Zimmerman AW. Immunitet, gliaceller och neuroinflammation vid autism. Int Rev Psychiatry. 2005;17(6):485-495.https://doi.org/10.1080/02646830500381930
[B78] Edmonson C, Ziats MN, Rennert OM. Altererat uttryck av gliamarkörer i autistisk postmortem prefrontala cortex och cerebellum. Mol Autism. 2014;5:3.https://doi.org/10.1186/2040-2392-5-3
[B79] Salter MW, Stevens B. Microglia emerge as central players in brain disease. Nat Med. 2017;23(9):1018-1027.https://doi.org/10.1038/nm.4397
[B80] Morgan JT, Chana G, Pardo CA, et al. Mikrogliamental aktivering och ökad mikrogliatäthet observerad i dorsolaterala prefrontala cortex vid autism. Biol Psychiatry. 2010;68(4):368-376.https://doi.org/10.1016/j.biopsych.2010.05.024
[B81] Suzuki K, Sugihara G, Ouchi Y, et al. Mikroaktivering hos unga vuxna med autism. JAMA Psychiatry. 2013;70(1):49-58.https://doi.org/10.1001/jamapsychiatry.2013.272
[B82] Zürcher NR, Loggia ML, Lawson R, et al. Ökad in vivo glial aktivering hos patienter med amyotrofisk lateralskleros: bedömd med [(11)C]-PBR28. Neuroimage Clin. 2015;7:409-414.https://doi.org/10.1016/j.nicl.2015.01.009
[B83] Schafer DP, Lehrman EK, Kautzman AG, et al. Microglia formar postnatal neurala kretsar på ett aktivitets- och komplementberoende sätt. Neuron. 2012;74(4):691-705.https://doi.org/10.1016/j.neuron.2012.03.026
[B84] Ashwood P, Krakowiak P, Hertz-Picciotto I, Hansen R, Pessah I, Van de Water J. Förhöjda plasmakitokiner vid autismdiagnoser ger bevis för immunologisk dysfunktion och är associerade med försämrat beteendemässigt utfall. Brain Behav Immun. 2011;25(1):40-45.https://doi.org/10.1016/j.bbi.2010.08.003
[B85] Li X, Chauhan A, Sheikh AM, et al. Förhöjt immunrespons i hjärnan hos autistiska patienter. J Neuroimmunol. 2009;207(1-2):111-116.https://doi.org/10.1016/j.jneuroim.2008.12.002
[B86] Smith SE, Li J, Garbett K, Mirnics K, Patterson PH. Maternal immune activation alters fetal brain development through interleukin-6. J Neurosci. 2007;27(40):10695-10702.https://doi.org/10.1523/JNEUROSCI.2178-07.2007
[B87] Choi GB, Yim YS, Wong H, med flera. Den modersbundna interleukin-17a-vägen hos möss främjar autism-liknande fenomen hos avkomman. Science. 2016;351(6276):933-939.https://doi.org/10.1126/science.aad0314
[B88] Stellwagen D, Malenka RC. Synaptic scaling mediated by glial TNF-alpha. Nature. 2006;440(7087):1054-1059.https://doi.org/10.1038/nature04671
[B89] Wei H, Chadman KK, McCloskey DP, m.fl. Förhöjt IL-6 i hjärnan orsakar obalanser i neuronala nätverk och medierar autism-liknande beteenden. Biochim Biophys Acta. 2012;1822(6):831-842.https://doi.org/10.1016/j.bbadis.2012.01.011
[B90] Verkhratsky A, Nedergaard M. Physiology of Astroglia. Physiol Rev. 2018;98(1):239-389.https://doi.org/10.1152/physrev.00042.2016
[B91] Laurence JA, Fatemi SH. Glial fibrillary acidic protein är förhöjt i cortex i överkäkshalvan, hjässloben och lillhjärnan hos autistiska individer. Cerebellum. 2005;4(3):206-210.https://doi.org/10.1080/14734220500208846
Knockout av glutamattransportörer avslöjar en stor roll för astroglial transport i excitotoxicitet och rensning av glutamat.https://doi.org/10.1016/s0896-6273(00 )80086-0
[B93] Sies H, Jones DP. Reactive oxygen species (ROS) som pleiotropa fysiologiska signaleringsagenter. Nat Rev Mol Cell Biol. 2020;21(7):363-383.https://doi.org/10.1038/s41580-020-0230-3
[B94] Frustaci A, Neri M, Cesario A, et al. Oxidativ stressrelaterade biomarkörer vid autism: systematisk översikt och meta-analyser. Free Radic Biol Med. 2012;52(10):2128-2141.https://doi.org/10.1016/j.freeradbiomed.2012.03.011
[B95] Bjørklund G, Meguid NA, El-Bana MA, et al. Oxidativ stress vid autismspektrumstörning. Mol Neurobiol. 2020;57(5):2314-2332.https://doi.org/10.1007/s12035-019-01742-2
[B96] James SJ, Cutler P, Melnyk S, et al. Metaboliska biomarkörer för ökad oxidativ stress och nedsatt metyleringskapacitet hos barn med autism. Am J Clin Nutr. 2004;80(6):1611-1617.https://doi.org/10.1093/ajcn/80.6.1611
[B97] Rose S, Melnyk S, Pavliv O, et al. Bevis på oxidativ skada och inflammation i samband med låg glutationredoxstatus i autismhjärnan. Transl Psychiatry. 2012;2:e134.https://doi.org/10.1038/tp.2012.61
[B98] Bowers K, Li Q, Bressler J, et al. Glutathione pathway gene variation and risk of autism spectrum disorders. J Neurodev Disord. 2011;3(2):132-143.https://doi.org/10.1007/s11689-011-9077-4
[B99] Rossignol DA, Frye RE. Mitochondrial dysfunction in autism spectrum disorders: a systematic review and meta-analysis. Mol Psychiatry. 2012;17(3):290-314.https://doi.org/10.1038/mp.2010.136
[B100] Giulivi C, Zhang YF, Omanska-Klusek A, m.fl. Mitokondriell dysfunktion vid autism. JAMA. 2010;304(21):2389-2396.https://doi.org/10.1001/jama.2010.1706
[B101] Duchen MR. Mitokondrier och kalcium: från signalsubstans till celldöd. J Physiol. 2000;529 Pt 1(Pt 1):57-68.https://doi.org/10.1111/j.1469-7793.2000.00057.x
[B102] Bambini-Junior V, Rodrigues L, Behr GA, et al. Djurmodell för autism inducerad av prenatal exponering för valproat: beteendeförändringar och leverparametrar. Brain Res. 2011;1408:8-16.https://doi.org/10.1016/j.brainres.2011.06.015
[B103] Ma Q. Roll av nrf2 vid oxidativ stress och toxicitet. Annu Rev Pharmacol Toxicol. 2013;53:401-426.https://doi.org/10.1146/annurev-pharmtox-011112-140320
[B104] Singh K, Connors SL, Macklin EA, et al. Sulforafanbehandling av autismspektrumstörning (ASD). Proc Natl Acad Sci U S A. 2014;111(43):15550-15555.https://doi.org/10.1073/pnas.1416940111
[B105] Bent S, Lawton B, Warren T, et al. Identifiering av urinmetaboliter som korrelerar med kliniska förbättringar hos barn med autism som behandlats med sulforafan från broccoli. Mol Autism. 2018;9:35.https://doi.org/10.1186/s13229-018-0218-4
[B106] Cryan JF, O'Riordan KJ, Cowan CSM, et al. The Microbiota-Gut-Brain Axis. Physiol Rev. 2019;99(4):1877-2013.https://doi.org/10.1152/physrev.00018.2018
[B107] Chaidez V, Hansen RL, Hertz-Picciotto I. Gastrointestinala problem hos barn med autism, utvecklingsförseningar eller typisk utveckling. J Autism Dev Disord. 2014;44(5):1117-1127.https://doi.org/10.1007/s10803-013-1973-x
[B108] Adams JB, Johansen LJ, Powell LD, Quig D, Rubin RA. Gastrointestinal flora och gastrointestinal status hos barn med autism – jämförelser med typiska barn och korrelation med autismens svårighetsgrad. BMC Gastroenterol. 2011;11:22.https://doi.org/10.1186/1471-230X-11-22
[B109] Xu M, Xu X, Li J, Li F. Association Between Gut Microbiota and Autism Spectrum Disorder: A Systematic Review and Meta-Analysis. Front Psychiatry. 2019;10:473.https://doi.org/10.3389/fpsyt.2019.00473
[B110] Liu F, Li J, Wu F, et al. Förändrad sammansättning och funktion av tarmmikrobiotan vid autismspektrumstörningar: en systematisk översikt. Transl Psychiatry. 2019;9(1):43.https://doi.org/10.1038/s41398-019-0389-6
[B111] Kang DW, Park JG, Ilhan ZE, et al. Minskad förekomst av Prevotella och andra fermenterare i tarmfloran hos autistiska barn. PLoS One. 2013;8(7):e68322.https://doi.org/10.1371/journal.pone.0068322
[B112] Stilling RM, van de Wouw M, Clarke G, Stanton C, Dinan TG, Cryan JF. Neurofarmakologin hos butyrat: Mikrobiota-tarm-hjärna-axelns basföda? Neurochem Int. 2016;99:110-132.https://doi.org/10.1016/j.neuint.2016.06.011
[B113] MacFabe DF, Cain DP, Rodriguez-Capote K, et al. Neurobiologiska effekter av intraventrikulär propionsyra hos råttor: möjlig roll för kortkedjiga fettsyror i patogenesen och egenskaperna hos autism-spektrumstörningar. Behav Brain Res. 2007;176(1):149-169.https://doi.org/10.1016/j.bbr.2006.07.025
[B114] Hsiao EY, McBride SW, Hsien S, et al. Mikrobiota modulerar beteendemässiga och fysiologiska abnormaliteter associerade med neuropsykiatriska funktionsnedsättningar. Cell. 2013;155(7):1451-1463.https://doi.org/10.1016/j.cell.2013.11.024
[B115] Bonaz B, Bazin T, Pellissier S. Vagusnerven vid gränsytan mellan mikrobiota-tarm-hjärna-axeln. Front Neurosci. 2018;12:49.https://doi.org/10.3389/fnins.2018.00049
[B116] Liu S, Hagiwara SI, Bhargava A. Tidiga livets motgångar, epigenetik och visceral hypersensitivitet. Neurogastroenterol Motil. 2017;29(9).https://doi.org/10.1111/nmo.13170
[B117] Zoghbi HY, Bear MF. Synaptic dysfunction in neurodevelopmental disorders associated with autism and intellectual disabilities. Cold Spring Harb Perspect Biol. 2012;4(3):a009886.https://doi.org/10.1101/cshperspect.a009886
[B118] Bourgeron T. Från den genetiska arkitekturen till synaptisk plasticitet vid autism. Nat Rev Neurosci. 2015;16(9):551-563.https://doi.org/10.1038/nrn3992
[B119] Südhof TC. Neuroliginer och neurexiner kopplar synaptisk funktion till kognitiv sjukdom. Nature. 2008;455(7215):903-911.https://doi.org/10.1038/nature07456
[B120] Jamain S, Quach H, Betancur C, et al. Mutationer i X-kromosombundna gener som kodar för neuroliginer NLGN3 och NLGN4 är associerade med autism. Nat Genet. 2003;34(1):27-29.https://doi.org/10.1038/ng1136
[B121] Tabuchi K, Blundell J, Etherton MR, et al. En mutation i neuroligin-3 som är kopplad till autism ökar hämmande synaptisk transmission hos möss. Science. 2007;318(5847):71-76.https://doi.org/10.1126/science.1146221
[B122] Jamain S, Radyushkin K, Hammerschmidt K, et al. Minskad social interaktion och ultraljudskommunikation i en musmodell av monogen ärftlig autism. Proc Natl Acad Sci U S A. 2008;105(5):1710-1715.https://doi.org/10.1073/pnas.0711555105
[B123] Grabrucker AM, Schmeisser MJ, Schoen M, Boeckers TM. Postsynaptiska ProSAP/Shank-ställningar i korseld av synaptopatier. Trends Cell Biol. 2011;21(10):594-603.https://doi.org/10.1016/j.tcb.2011.07.003
[B124] Phelan K, McDermid HE. Phelan-McDermid syndrom (22q13.3 deletionssyndrom). Mol Syndromol. 2012;2(3-5):186-201.https://doi.org/10.1159/000334260
[B125] Peça J, Feliciano C, Ting JT, et al. Shank3-mutantmöss uppvisar autismeliknande beteenden och dysfunktion i striatum. Nature. 2011;472(7344):437-442.https://doi.org/10.1038/nature09965
[B126] Won H, Lee HR, Gee HY, et al. Autistiskt liknande socialt beteende hos Shank2-muterade möss förbättrades genom att återställa NMDA-receptorns funktion. Nature. 2012;486(7402):261-265.https://doi.org/10.1038/nature11208
[B127] Baudouin SJ, Gaudias J, Gerharz S, et al. Delad synaptisk patofysiologi i syndromala och icke-syndromala gnagarmdeller av autism. Science. 2012;338(6103):128-132.https://doi.org/10.1126/science.1224159
[B128] Huber KM, Gallagher SM, Warren ST, Bear MF. Altered synaptic plasticity in a mouse model of fragile X mental retardation. Proc Natl Acad Sci U S A. 2002;99(11):7746-7750.https://doi.org/10.1073/pnas.122205699
[B129] Ecker C, Bookheimer SY, Murphy DG. Neuroimaging i autismspektrumtillstånd: hjärnans struktur och funktion genom hela livet. Lancet Neurol. 2015;14(11):1121-1134.https://doi.org/10.1016/S1474-4422(15 )00050-2
[B130] Hazlett HC, Gu H, Munsell BC, et al. Tidig hjärnans utveckling hos spädbarn med hög risk för autism. Nature. 2017;542(7641):348-351.https://doi.org/10.1038/nature21369
[B131] Miller EK, Cohen JD. En integrerande teori om prefrontala cortex funktion. Annu Rev Neurosci. 2001;24:167-202.https://doi.org/10.1146/annurev.neuro.24.1.167
[B132] Castelli F, Frith C, Happé F, Frith U. Autism, Asperger syndrom och hjärnmekanismer för tillskrivande av mentala tillstånd till animerade former. Brain. 2002;125(Pt 8):1839-1849.https://doi.org/10.1093/brain/awf189
[B133] Just MA, Cherkassky VL, Keller TA, Kana RK, Minshew NJ. Funktionell och anatomisk kortikal underkonnektivitet vid autism: bevis från en fMRI-studie av en exekutiv funktion-uppgift och corpus callosum morfometri. Cereb Cortex. 2007;17(4):951-961.https://doi.org/10.1093/cercor/bhl006
[B134] Casanova MF, Buxhoeveden DP, Switala AE, Roy E. Minikolumnär patologi vid autism. Neurology. 2002;58(3):428–432.https://doi.org/10.1212/wnl.58.3.428
[B135] Adolphs R. The social brain: neural basis of social knowledge. Annu Rev Psychol. 2009;60:693-716.https://doi.org/10.1146/annurev.psych.60.110707.163514
[B136] Nordahl CW, Scholz R, Yang X, et al. Ökad tillväxttakt av amygdala hos barn i åldrarna 2 till 4 år med autism-spektrumtillstånd: en longitudinell studie. Arch Gen Psychiatry. 2012;69(1):53-61.https://doi.org/10.1001/archgenpsychiatry.2011.145
[B137] Kleinhans NM, Richards T, Sterling L, et al. Abnormell funktionell konnektivitet vid autismspektrumtillstånd under ansiktsbearbetning. Brain. 2008;131(Pt 4):1000-1012.https://doi.org/10.1093/brain/awm334
[B138] Monk CS, Weng SJ, Wiggins JL, et al. Neural circuitry of emotional face processing in autism spectrum disorders. J Psychiatry Neurosci. 2010;35(2):105-114.https://doi.org/10.1503/jpn.090085
[B139] Schmahmann JD. Lilla hjärnan och kognition. Neurosci Lett. 2019;688:62-75.https://doi.org/10.1016/j.neulet.2018.07.005
[B140] Fatemi SH, Aldinger KA, Ashwood P, m.fl. Konsensuspapper: patologisk roll av lillhjärnan vid autism. Cerebellum. 2012;11(3):777-807.https://doi.org/10.1007/s12311-012-0355-9
[B141] Courchesne E, Yeung-Courchesne R, Press GA, Hesselink JR, Jernigan TL. Hypoplasi av cerebellära vermis lobuli VI och VII vid autism. N Engl J Med. 1988;318(21):1349-1354.https://doi.org/10.1056/NEJM198805263182102
[B142] Wang SS, Kloth AD, Badura A. Lilla hjärnan, känsliga perioder och autism. Neuron. 2014;83(3):518-532.https://doi.org/10.1016/j.neuron.2014.07.016
[B143] Squire LR, Stark CE, Clark RE. Mediala temporala loben. Annu Rev Neurosci. 2004;27:279-306.https://doi.org/10.1146/annurev.neuro.27.070203.144130
[B144] Schumann CM, Hamstra J, Goodlin-Jones BL, et al. Amygdala är förstorad hos barn men inte hos ungdomar med autism; hippocampus är förstorad i alla åldrar. J Neurosci. 2004;24(28):6392-6401.https://doi.org/10.1523/JNEUROSCI.1297-04.2004
[B145] Gaigg SB, Bowler DM. Fritt återgivande och glömska av känslomässigt stimulerande ord vid autism-spektrumtillstånd. Neuropsychologia. 2008;46(9):2336-2343.https://doi.org/10.1016/j.neuropsychologia.2008.03.008
[B146] Graybiel AM. Basala ganglierna. Curr Biol. 2000;10(14):R509-R511.https://doi.org/10.1016/s0960-9822(00 )00593-5
[B147] Langen M, Durston S, Staal WG, Palmen SJ, van Engeland H. Caudatuskärnan är förstorad hos högfungerande, medicinfria individer med autism. Biol Psychiatry. 2007;62(3):262-266.https://doi.org/10.1016/j.biopsych.2006.09.040
[B148] Scott-Van Zeeland AA, Dapretto M, Ghahremani DG, Poldrack RA, Bookheimer SY. Belöningsprocessande vid autism. Autism Res. 2010;3(2):53-67.https://doi.org/10.1002/aur.122
[B149] Gershenzon J, Dudareva N. Terpeners naturliga produkters funktion i den naturliga världen. Nat Chem Biol. 2007;3(7):408-414.https://doi.org/10.1038/nchembio.2007.5
[B150] Russo EB. Taming THC: potential cannabis synergy and phytocannabinoid-terpenoid entourage effects. Br J Pharmacol. 2011;163(7):1344-1364.https://doi.org/10.1111/j.1476-5381.2011.01238.x
[B151] Aprotosoaie AC, Hăncianu M, Costache II, Miron A. Linalool: en översikt av en viktig doftmolekyl med värdefulla biologiska egenskaper. Flavour Fragr J. 2014;29(4):193-219.https://doi.org/10.1002/ffj.3197
[B152] Harada H, Kashiwadani H, Kanmura Y, Kuwaki T. Linanolldoftinducerade anxiolytiska effekter hos möss. Front Behav Neurosci. 2018;12:241.https://doi.org/10.3389/fnbeh.2018.00241
[B153] Elisabetsky E, Brum LF, Souza DO. Antikonvulsiva egenskaper hos linalool i glutamatrelaterade epileptiska modellfall. Phytomedicine. 1999;6(2):107-113.https://doi.org/10.1016/S0944-7113(99 )80044-0
[B154] Linck VM, da Silva AL, Figueiró M, et al. Effekter av inhalerad linalool på ångest, social interaktion och aggressivt beteende hos möss. Phytomedicine. 2010;17(8-9):679-683.https://doi.org/10.1016/j.phymed.2009.10.002
[B155] Takahashi M, Satou T, Ohashi M, Hayashi S, Sadamoto K, Koike K. Interspecies-jämförelse av kemisk sammansättning och ångestdämpande effekter av lavendeloljor vid inandning. Nat Prod Commun. 2011;6(11):1769-1774.https://journals.sagepub.com/doi/10.1177/1934578X1100601148
[B156] Peana AT, D’Aquila PS, Panin F, Serra G, Pippia P, Moretti MD. Antiinflammatorisk aktivitet hos linalool och linalylacetat som beståndsdelar i eteriska oljor. Phytomedicine. 2002;9(8):721-726.https://doi.org/10.1078/094471102321621322
[B157] Sabogal-Guáqueta AM, Posada-Duque R, Cortes NC, Arias-Londoño JD, Cardona-Gómez GP. Förändringar i den hippokampala och perifera fosfolipidprofilen är associerade med kännetecken för neurodegeneration i en långvarig modell av global cerebral ischemi: Hämning av Linalool. Neuropharmacology. 2018;135:555-571.https://doi.org/10.1016/j.neuropharm.2018.04.015
[B158] Jäger W, Buchbauer G, Jirovetz L, Fritzer M. Perkutän absorption av lavendelolja från en massageolja. J Soc Cosmet Chem. 1992;43:49-54.https://www.researchgate.net/publication/232711524_Percutaneous_absorption_of_lavender_oil_from_a_massage_oil
[B159] Buchbauer G. Biologiska aktiviteter hos eteriska oljor. I: Baser KHC, Buchbauer G, red. Handbook of Essential Oils: Science, Technology, and Applications. 2:a utgåvan. CRC Press; 2015:235-280.https://www.routledge.com/Handbook-of-Essential-Oils-Science-Technology-and-Applications/Baser-Buchbauer/p/book/9781032046419
[B160] Gertsch J, Leonti M, Raduner S, et al. Beta-karyofyllen är en kostrelaterad cannabinoid. Proc Natl Acad Sci U S A. 2008;105(26):9099-9104.https://doi.org/10.1073/pnas.0803601105
[B161] Bahi A, Al Mansouri S, Al Memari E, Al Ameri M, Nurulain SM, Ojha S. β-Caryophyllene, en CB2-receptoragonist, producerar multipla beteendeförändringar relevanta för ångest och depression hos möss. Physiol Behav. 2014;135:119-124.https://doi.org/10.1016/j.physbeh.2014.06.003
[B162] Atwood BK, Mackie K. CB2: en kannabinoidreceptor med en identitetskris. Br J Pharmacol. 2010;160(3):467-479.https://doi.org/10.1111/j.1476-5381.2010.00729.x
[B163] Klauke AL, Racz I, Pradier B, et al. Den cannabinoida CB₂-receptorselektiva fytocannabinoiden beta-karyofyllen utövar smärtlindrande effekter i musmodeller av inflammatorisk och neuropatisk smärta. Eur Neuropsychopharmacol. 2014;24(4):608-620.https://doi.org/10.1016/j.euroneuro.2013.10.008
[B164] Cheng Y, Dong Z, Liu S. β-Caryophyllene lindrar Alzheimer-liknande fenotyp hos APP/PS1-möss genom CB2-receptoraktivering och PPARγ-vägen. Pharmacology. 2014;94(1-2):1-12.https://doi.org/10.1159/000362689
[B165] Askari VR, Shafiee-Nick R. Lovande neuroprotektiva effekter av β-karyofyllen mot LPS-inducerad oligodendrocyt-toxicitet: En mekanistisk studie. Biochem Pharmacol. 2019;159:154-171.https://doi.org/10.1016/j.bcp.2018.12.001
[B166] Fontes LBA, Dias D, Aarestrup BJ, et al. β-Caryophyllene lindrar utvecklingen av experimentell autoimmun encefalomyelit hos C57BL/6-möss. Biomed Pharmacother. 2017;91:257-264.https://doi.org/10.1016/j.biopha.2017.04.092
[B167] Alberti TB, Barbosa WL, Vieira JL, Raposo NR, Dutra RC. (-)-β-Caryophyllene, ett CB2-receptor-selektivt fytocannabinoid, undertrycker motorisk paralys och neuroinflammation i en murin modell av multipel skleros. Int J Mol Sci. 2017;18(4):691.https://doi.org/10.3390/ijms18040691
[B168] Galdino PM, Nascimento MV, Florentino IF, et al. Den ångestdämpande effekten av en eterisk olja från Spiranthera odoratissima A. St. Hil. blad och dess huvudsakliga komponent, β-karyofyllen, hos hanmöss. Prog Neuropsychopharmacol Biol Psychiatry. 2012;38(2):276-284.https://doi.org/10.1016/j.pnpbp.2012.04.012
[B169] Horváth B, Mukhopadhyay P, Kechrid M, et al. β-Caryofyllen lindrar cisplatin-inducerad nefrotoxicitet på ett cannabinoid 2-receptorbundet sätt. Free Radic Biol Med. 2012;52(8):1325-1333.https://doi.org/10.1016/j.freeradbiomed.2012.01.014
Fidyt K, Fiedorowicz A, Strządała L, Szumny A. β-karyofyllen och β-karyofyllenoxid – naturliga föreningar med antikancer- och smärtstillande egenskaper. Cancer Med. 2016;5(10):3007-3017.https://doi.org/10.1002/cam4.816
[B171] Sharma C, Al Kaabi JM, Nurulain SM, Goyal SN, Kamal MA, Ojha S. Polyfarmakologiska egenskaper och terapeutisk potential hos β-karyofyllen: en kostförekomst av fytokannabinoider med farmaceutisk potential. Curr Pharm Des. 2016;22(21):3237-3264.https://doi.org/10.2174/1381612822666160311115226
[B172] Ammon HP. Boswellinsyror och deras roll vid kroniska inflammatoriska sjukdomar. Adv Exp Med Biol. 2016;928:291-327.https://doi.org/10.1007/978-3-319-41334-1_13
[B173] Catanzaro D, Rancan S, Orso G, et al. Boswellia serrata bevarar tarmens epitelbarriär från oxidativ och inflammatorisk skada. PLoS One. 2015;10(5):e0125375.https://doi.org/10.1371/journal.pone.0125375
[B174] Khoshnazar M, Bigdeli MR, Parvardeh S, Pouriran R. Dämpande effekt av α-pinen på neurobeteendemässiga brister, oxidativ skada och inflammatorisk respons efter fokalt ischemisk stroke hos råtta. J Pharm Pharmacol. 2019;71(11):1725-1733.https://doi.org/10.1111/jphp.13164
[B175] Ryter SW, Alam J, Choi AM. Hemoxygenas-1 / kolmonoxid: från grundvetenskap till terapeutiska tillämpningar. Physiol Rev. 2006;86(2):583-650.https://doi.org/10.1152/physrev.00011.2005
[B176] Umar S, Umar K, Sarwar AH, et al. Boswellia serrata-extrakt dämpar inflammatoriska medlare och oxidativ stress vid kollageninducerad artrit. Phytomedicine. 2014;21(6):847-856.https://doi.org/10.1016/j.phymed.2014.02.001
[B177] Abdel-Tawab M, Werz O, Schubert-Zsilavecz M. Boswellia serrata: en övergripande bedömning av in vitro, prekliniska, farmakokinetiska och kliniska data. Clin Pharmacokinet. 2011;50(6):349-369.https://doi.org/10.2165/11586800-000000000-00000
[B178] Hosseini M, Hadjzadeh MA, Derakhshan M, et al. De positiva effekterna av olibanum på minnesunderskott inducerat av hypotyreos hos vuxna råttor testade i Morris vatten labyrint. Arch Pharm Res. 2010;33(3):463-468.https://doi.org/10.1007/s12272-010-0316-6
[B179] Safayhi H, Mack T, Sabieraj J, Anazodo MI, Subramanian LR, Ammon HP. Boswellinsyror: nya, specifika, icke redox-hämmare av 5-lipoxygenas. J Pharmacol Exp Ther. 1992;261(3):1143-1146.https://www.sciencedirect.com/science/article/abs/pii/S0022356525111300
[B180] Werz O. 5-lipoxygenas: cellulär biologi och molekylär farmakologi. Curr Drug Targets Inflammation Allergy. 2002;1(1):23-44.https://doi.org/10.2174/1568010023344959
[B181] Al-Harrasi A, Ali L, Hussain J, et al. Analgetiska effekter av råextrakt och fraktioner av omanisk rökelse från traditionella medicinalväxter Boswellia sacra på djurmodeller. Asian Pac J Trop Med. 2014;7S1:S485-S490.https://doi.org/10.1016/S1995-7645(14 )60279-0
[B182] Cimanga K, Kambu K, Tona L, et al. Correlation between chemical composition and antibacterial activity of essential oils of some aromatic medicinal plants growing in the Democratic Republic of Congo. J Ethnopharmacol. 2002;79(2):213-220.https://doi.org/10.1016/s0378-8741(01 )00384-1
[B183] Satou T, Kasuya H, Maeda K, Koike K. Daglig inandning av α-pinen hos möss: effekter på beteende och organackumulering. Phytother Res. 2014;28(9):1284-1287.https://doi.org/10.1002/ptr.5105
[B184] Satou T, Murakami S, Matsuura M, Hayashi S, Koike K. Ångestdämpande effekt och vävnadsfördelning av inhalerad eterisk olja från Alpinia zerumbet hos möss. Nat Prod Commun. 2010;5(1):143-146.https://journals.sagepub.com/doi/10.1177/1934578X1000500133
[B185] Juergens UR, Dethlefsen U, Steinkamp G, Gillissen A, Repges R, Vetter H. Antiinflammatorisk aktivitet av 1,8-kineol (eukalyptol) vid bronkialastma: en dubbelblind placebokontrollerad studie. Respir Med. 2003;97(3):250-256.https://doi.org/10.1053/rmed.2003.1432
[B186] Santos FA, Rao VS. Antiinflammatoriska och antinociceptiva effekter av 1,8-kineol en terpenoidoxid som finns i många eteriska oljor från växter. Phytother Res. 2000;14(4):240-244.https://doi.org/10.1002/1099-1573(200006 )14:43.0.co;2-x
[B187] Tisserand R, Young R. Essential Oil Safety: A Guide for Health Care Professionals. 2nd ed. Churchill Livingstone; 2014.https://www.elsevier.com/books/essential-oil-safety/tisserand/978-0-7020-4606-4
[B188] Worth H, Schacher C, Dethlefsen U. Concomitant therapy with Cineole (Eucalyptole) reductions exacserbationer vid KOL: en placebokontrollerad dubbelblind studie. Respir Res. 2009;10:69.https://doi.org/10.1186/1465-9921-10-69
[B189] Salehi B, Upadhyay S, Erdogan Orhan I, et al. Terapeutisk potential för α- och β-Pinene: En mirakelgåva från naturen. Biomolekylär. 2019;9(11):738.https://doi.org/10.3390/biom9110738
[B190] Guzmán-Gutiérrez SL, Gómez-Cansino R, García-Zebadúa JC, Jiménez-Pérez NC, Reyes-Chilpa R. Antidepressiv aktivitet av eterisk olja från Litsea glaucescens: identifiering av β-pinen och linalool som aktiva principer. J Ethnopharmacol. 2012;143(2):673-679.https://doi.org/10.1016/j.jep.2012.07.026
[B191] Yang H, Woo J, Pae AN, et al. α-Pinene, en viktig beståndsdel i talloljor, förbättrar sömn utan snabb ögonrörelse hos möss genom GABAA-benzodiazepinreceptorer. Mol Pharmacol. 2016;90(5):530-539.https://doi.org/10.1124/mol.116.105080
[B192] Kim DS, Lee HJ, Jeon YD, et al. Alfa-pinen uppvisar antiinflammatorisk aktivitet genom suppression av MAPKs och NF-κB-vägen i musperitoneala makrofager. Am J Chin Med. 2015;43(4):731-742.https://doi.org/10.1142/S0192415X15500457
[B193] Porres-Martínez M, González-Burgos E, Carretero ME, Gómez-Serranillos MP. Skyddande egenskaper hos Salvia lavandulifolia Vahl. eterisk olja mot oxidativ stressinducerad neuronal skada. Food Chem Toxicol. 2015;80:154-162.https://doi.org/10.1016/j.fct.2015.03.002
[B194] Miyazawa M, Watanabe H, Kameoka H. Inhibering av acetylkolinesterasaktivitet av monoterpenoider med ett p-menthanskelett. J Agric Food Chem. 1997;45(3):677-679.https://doi.org/10.1021/jf960398b
[B195] Deutsch SI, Burket JA, Katz E. Bidrar subtil störning av neuronal migration till schizofreni och andra neurologiska utvecklingsstörningar? Potentiella genetiska mekanismer med möjliga behandlingsimplikationer. Eur Neuropsychopharmacol. 2010;20(5):281-287.https://doi.org/10.1016/j.euroneuro.2010.02.005
[B196] Tan LT, Lee LH, Yin WF, et al. Traditionella användningsområden, fytokemi och bioaktiviteter hos Cananga odorata (Ylang-Ylang). Evid Based Complement Alternat Med. 2015;2015:896314.https://doi.org/10.1155/2015/896314
[B197] Jung YH, Ha TM, Ahn J, et al. Inandning av Cananga odorata eterisk olja förbättrade social interaktion och ångestliknande beteenden hos valproinsyrainducerade autismspektrumstörningsråttor. Näringsämnen. 2021;13(11):3804.https://doi.org/10.3390/nu13113804
[B198] Sinha Y, Silove N, Wheeler D, Williams K. Auditory integration training and other sound therapies for autism spectrum disorders: a systematic review. Arch Dis Child. 2006;91(12):1018-1022.https://doi.org/10.1136/adc.2006.094649
[B199] Cullen-Powell LA, Barlow JH, Cushway D. Utforska en massageintervention för föräldrar och deras barn med autism: konsekvenserna för bindning och anknytning. J Child Health Care. 2005;9(4):245-255.https://doi.org/10.1177/1367493505056479
[B200] Buckle J. Klinisk aromaterapi: Eteriska oljor inom hälso- och sjukvården. 3:e upplagan, Churchill Livingstone; 2014.https://www.elsevier.com/books/clinical-aromatherapy/buckle/978-0-7020-5440-3
[B201] Saiyudthong S, Marsden CA. Akuta effekter av bergamottolja på ångestrelaterat beteende och kortikosteronnivå hos råttor. Phytother Res. 2011;25(6):858-862.https://doi.org/10.1002/ptr.3325
[B202] Navarra M, Mannucci C, Delbò M, Calapai G. Citrus bergamia eterisk olja: från grundforskning till klinisk tillämpning. Front Pharmacol. 2015;6:36.https://doi.org/10.3389/fphar.2015.00036
[B203] Efferth T, Oesch F. Antiinflammatoriska och antikreftaktiviteter av frankincense: Mål, behandlingar och toxiciteter. Semin Cancer Biol. 2022;80:39-57.https://doi.org/10.1016/j.semcancer.2020.01.015
[B204] Siddiqui MZ. Boswellia serrata, ett potentiellt antiinflammatoriskt medel: en översikt. Indian J Pharm Sci. 2011;73(3):255-261.https://doi.org/10.4103/0250-474X.93507
[B205] Hamidpour R, Hamidpour S, Hamidpour M, Shahlari M. Frankincense (rǔ xiāng; Boswellia-arter): Från urvalet av traditionella tillämpningar till den nya fytoterapin för förebyggande och behandling av allvarliga sjukdomar. J Tradit Complement Med. 2013;3(4):221-226.https://doi.org/10.4103/2225-4110.119723
[B206] Moussaieff A, Mechoulam R. Boswellia resin: från religiösa ceremonier till medicinsk användning; en genomgång av in-vitro, in-vivo och kliniska prövningar. J Pharm Pharmacol. 2009;61(10):1281-1293.https://doi.org/10.1211/jpp/61.10.0003
[B207] Bakkali F, Averbeck S, Averbeck D, Idaomar M. Biological effects of essential oils–a review. Food Chem Toxicol. 2008;46(2):446-475.https://doi.org/10.1016/j.fct.2007.09.106
[B208] Setzer WN. Eteriska oljor och ångestdämpande aromaterapi. Nat Prod Commun. 2009;4(9):1305-1316.https://pubmed.ncbi.nlm.nih.gov/19831048/
[B209] Adams RP. Identification of Essential Oil Components by Gas Chromatography/Mass Spectrometry. 4:e uppl. Allured Publishing; 2007.https://www.allured.com/identification-of-essential-oil-components-by-gas-chromatographymass-spectrometry-4th-edition
[B210] Dayawansa S, Umeno K, Takakura H, et al. Autonoma svar vid inandning av naturlig cedrol-doft hos människor. Auton Neurosci. 2003;108(1-2):79-86.https://doi.org/10.1016/j.autneu.2003.08.002
[B211] Cheaha D, Reakkamnuan C, Nukitram J, et al. Modifiering av sömn-vakna och elektroencefalogram inducerad av inhalation av eterisk olja från vetiver. J Intercult Ethnopharmacol. 2016;5(1):72-78.https://doi.org/10.5455/jice.20160208050736
[B212] Friedmann T. Fri från ADHD: Ett läkemedelsfritt tillvägagångssätt för behandling av barn med ADHD. Lotus Press; 2000.https://www.researchgate.net/publication/265127532_Attention_Deficit_and_Hyperactivity_Disorder_ADHD
[B213] Weyerstahl P, Marschall H, Weirauch M, Thefeld K, Surburg H. Constituents of commercial vetiver oils. Flavour Fragr J. 2000;15(3):153-173.https://doi.org/10.1002/1099-1026(200005/06 )15:33.0.CO;2-V
[B214] Pripdeevech P, Chumpolsri W, Suttiarporn P, Wongpornchai S. Den kemiska sammansättningen och antioxidantaktiviteterna hos basilika från Thailand med hjälp av retentionindex och omfattande tvådimensionell gaskromatografi. J Serb Chem Soc. 2010;75(11):1503-1513.https://doi.org/10.2298/JSC100203125P
[B215] Hongratanaworakit T, Buchbauer G. Avslappnande effekt av ylang ylang-olja på människor efter transdermalt upptag. Phytother Res. 2006;20(9):758-763.https://doi.org/10.1002/ptr.1950
[B216] Moss M, Hewitt S, Moss L, Wesnes K. Modulation av kognitiv prestation och humör genom dofter av pepparmynta och ylang-ylang. Int J Neurosci. 2008;118(1):59-77.https://doi.org/10.1080/00207450601042094
[B217] Komiya M, Takeuchi T, Harada E. Lemonoljaånga ger en antistresseffekt genom att modulera 5-HT- och DA-aktiviteten hos möss. Behav Brain Res. 2006;172(2):240-249.https://doi.org/10.1016/j.bbr.2006.05.006
[B218] Venturi GB. Experimentella undersökningar angående principen för den laterala kommunikationen av rörelse i vätskor. Phil Trans R Soc Lond. 1797;87:243-260.https://royalsocietypublishing.org/doi/10.1098/rstl.1799.0005
Buckle J. Användning av aromaterapi som komplementär behandling vid kronisk smärta. Altern Ther Health Med. 1999;5(5):42-51.https://pubmed.ncbi.nlm.nih.gov/10450129/
[B220] Hongratanaworakit T. Aromaterapeutiska effekter av massage med eteriska oljor blandade på människor. Nat Prod Commun. 2011;6(8):1199-1204.https://www.researchgate.net/publication/51647617_Aroma-therapeutic_Effects_of_Massage_Blended_Essential_Oils_on_Humans
[B221] White FM. Strömningsmekanik. 8:e uppl. McGraw-Hill Education; 2015.https://www.mheducation.com/highered/product/fluid-mechanics-white/M9781260258318.html
[B222] Haze S, Sakai K, Gozu Y. Effekter av doftinhalation på sympatisk aktivitet hos normala vuxna. Jpn J Pharmacol. 2002;90(3):247-253.https://doi.org/10.1254/jjp.90.247
[B223] Herz RS. Aromaterapins fakta och fiktion: en vetenskaplig analys av lukteffekter på humör, fysiologi och beteende. Int J Neurosci. 2009;119(2):263-290.https://doi.org/10.1080/00207450802333953
[B224] Turek C, Stintzing FC. Stabilitet hos eteriska oljor: en översikt. Compr Rev Food Sci Food Saf. 2013;12(1):40-53.https://doi.org/10.1111/1541-4337.12006
[B225] Buckle J. Klinisk aromaterapi inom omvårdnad. 2:a utgåvan. Holistic Nursing Practice; 2001.https://journals.lww.com/holistic-nursing-practice/citation/2001/04000/clinical_aromatherapy_in_nursing.15.aspx
[B226] Labiris NR, Dolovich MB. Läkemedelsleverans till lungorna. Del I: fysiologiska faktorer som påverkar den terapeutiska effekten av inhalerade läkemedel. Br J Clin Pharmacol. 2003;56(6):588-599.https://doi.org/10.1046/j.1365-2125.2003.01892.x
[B227] Price S, Price L. Aromatherapy for Health Professionals. 4:e upplagan. Churchill Livingstone; 2011.https://www.elsevier.com/books/aromatherapy-for-health-professionals/price/978-0-7020-3151-0
[B228] Battaglia S. The Complete Guide to Aromatherapy. 3:e uppl. Black Pepper Creative; 2018.https://www.blackpeppercreative.com.au/the-complete-guide-to-aromatherapy
[B229] Posadzki P, Alotaibi A, Ernst E. Biverkningar av aromaterapi: en systematisk översikt av fallrapporter och fallserier. Int J Risk Saf Med. 2012;24(3):147-161.https://doi.org/10.3233/JRS-2012-0568
[B230] Dunn C, Sleep J, Collett D. Sensing an improvement: an experimental study to evaluate the use of aromatherapy, massage and periods of rest in an intensive care unit. J Adv Nurs. 1995;21(1):34-40.https://doi.org/10.1046/j.1365-2648.1995.21010034.x
[B231] Bowles EJ. The Chemistry of Aromatherapeutic Oils. 3:e uppl. Allen & Unwin; 2003.https://www.allenandunwin.com/browse/books/general-books/health-wellbeing/The-Chemistry-of-Aromatherapeutic-Oils-E-Joy-Bowles-9781741140217
[B232] Schnaubelt K. Advanced Aromatherapy: The Science of Essential Oil Therapy. Healing Arts Press; 1998.https://www.healingartspressbooks.com/advanced-aromatherapy
[B233] Schaaf RC, Lane AE. Mot ett bästa praxisprotokoll för bedömning av sensoriska drag vid ASD. J Autism Dev Disord. 2015;45(5):1380-1395.https://doi.org/10.1007/s10803-014-2299-z
[B234] Wolkoff P, Wilkins CK, Clausen PA, Nielsen GD. Organiska föreningar i kontorsmiljöer – sensorisk irritation, lukt, mätningar och rollen av reaktiv kemi. Indoor Air. 2006;16(1):7-19.https://doi.org/10.1111/j.1600-0668.2005.00393.x
[B235] Dalton P. Olfactory adaptation och korsanpassning. I: Doty RL, red. Handbook of Olfaction and Gustation. 3:e uppl. Wiley-Blackwell; 2015:443-459.https://www.wiley.com/en-us/Handbook+of+Olfaction+and+Gustation%2C+3rd+Edition-p-9780470455951
[B236] Cain WS. Att känna med näsan: nycklar till identifiering av lukt. Science. 1979;203(4379):467-470.https://doi.org/10.1126/science.760202
[B237] Kern JK, Trivedi MH, Garver CR, et al. Mönstret av sensoriska bearbetningsavvikelser vid autism. Autism. 2006;10(5):480-494.https://doi.org/10.1177/1362361306066564
[B238] Lehrner J, Marwinski G, Lehr S, Johren P, Deecke L. Omgivande dofter av apelsin och lavendel minskar ångest och förbättrar humöret på en tandläkarmottagning. Physiol Behav. 2005;86(1-2):92-95.https://doi.org/10.1016/j.physbeh.2005.06.031
[B239] Cavanagh HM, Wilkinson JM. Biologiska aktiviteter av lavendel eterisk olja. Phytother Res. 2002;16(4):301-308.https://doi.org/10.1002/ptr.1103
[B240] Diego MA, Jones NA, Field T, et al. Aromaterapi påverkar positivt humör, EEG-mönster för vakenhet och matematiska beräkningar. Int J Neurosci. 1998;96(3-4):217-224.https://doi.org/10.3109/00207459808986469
[B241] Field T, Diego M, Hernandez-Reif M, Cisneros W, Feijo L, Vera Y, Gil K, Grina D, Claire He Q. Effekter av rengöringsgel med lavendeldoft på avslappning. Int J Neurosci. 2005;115(2):207-222.https://doi.org/10.1080/00207450590519175
[B242] Goel N, Kim H, Lao RP. Ett luktintryck modifierar nattlig sömn hos unga män och kvinnor. Chronobiol Int. 2005;22(5):889-904.https://doi.org/10.1080/07420520500263276
Ashburner J, Ziviani J, Rodger S. Sensory processing and classroom emotional, behavioral, and educational outcomes in children with autism spectrum disorder. Am J Occup Ther. 2008;62(5):564-573.https://doi.org/10.5014/ajot.62.5.564
[B244] Tomchek SD, Dunn W. Sensorisk bearbetning hos barn med och utan autism: en jämförande studie med hjälp av det korta sensoriska profilen. Am J Occup Ther. 2007;61(2):190-200.https://doi.org/10.5014/ajot.61.2.190
[B245] Ben-Sasson A, Hen L, Fluss R, Cermak SA, Engel-Yeger B, Gal E. En metaanalys av sensoriska moduleringssymptom hos individer med autism spektrumstörningar. J Autism Dev Disord. 2009;39(1):1-11.https://doi.org/10.1007/s10803-008-0593-3
[B246] Leekam SR, Nieto C, Libby SJ, Wing L, Gould J. Describing the sensory abnormalities of children and adults with autism. J Autism Dev Disord. 2007;37(5):894-910.https://doi.org/10.1007/s10803-006-0218-7
[B247] Baranek GT, David FJ, Poe MD, Stone WL, Watson LR. Sensory Experiences Questionnaire: discriminating sensory features in young children with autism, developmental delays, and typical development. J Child Psychol Psychiatry. 2006;47(6):591-601.https://doi.org/10.1111/j.1469-7610.2005.01546.x
[B248] Herz RS. Odörframkallade minnens roll för psykologisk och fysiologisk hälsa. Brain Sci. 2016;6(3):22.https://doi.org/10.3390/brainsci6030022
[B249] Mesibov GB, Shea V, Schopler E. The TEACCH Approach to Autism Spectrum Disorders. Springer; 2004.https://link.springer.com/book/10.1007/978-0-387-23855-2
[B250] Oono IP, Honey EJ, McConachie H. Föräldramedierad tidig intervention för små barn med autism (ASD). Cochrane Database Syst Rev. 2013;(4):CD009774.https://doi.org/10.1002/14651858.CD009774.pub2
[B251] Marriott PJ, Shellie R, Cornwell C. Gas chromatographic technologies for the analysis of essential oils. J Chromatogr A. 2001;936(1-2):1-22.https://doi.org/10.1016/s0021-9673(01 )01314-0
[B252] Sparkman OD, Penton Z, Kitson FG. Gas Chromatography and Mass Spectrometry: A Practical Guide. 2:a uppl. Academic Press; 2011.https://www.sciencedirect.com/book/9780123736284/gas-chromatography-and-mass-spectrometry
[B253] ISO 4720:2009. Eteriska oljor – Nomenklatur. International Organization for Standardization; 2009.https://www.iso.org/standard/41076.html
[B254] ISO 11024-1:1998. Eteriska oljor – Allmän vägledning om kromatografiska profiler – Del 1: Framställning av kromatografiska profiler för presentation i standarder. International Organization for Standardization; 1998.https://www.iso.org/standard/19053.html
[B255] Dhifi W, Bellili S, Jazi S, Bahloul N, Mnif W. Essential Oils’ Chemical Characterization and Investigation of Some Biological Activities: A Critical Review. Medicines (Basel). 2016;3(4):25.https://doi.org/10.3390/medicines3040025
[B256] Lis-Balchin M. Aromatherapy Science: A Guide for Healthcare Professionals. Pharmaceutical Press; 2006.https://www.pharmpress.com/product/9780853696230/aromatherapy-science
[B257] Schnaubelt K. Medicinsk aromaterapi: Läkning med eteriska oljor. Frog Books; 1999.https://www.innertraditions.com/books/medical-aromatherapy
[B258] Tisserand R. The Art of Aromatherapy. Healing Arts Press; 1977.https://www.innertraditions.com/books/the-art-of-aromatherapy
[B259] Pauli A. Antimikrobiella egenskaper hos eteriska oljekomponenter. Int J Aromather. 2001;11(3):126-133.https://doi.org/10.1016/S0962-4562(01 )80048-5
[B260] Europeiska farmakopénämnden. Europeiska farmakopén. 10:e uppl. Europarådet; 2019.https://www.edqm.eu/en/european-pharmacopoeia-ph-eur-11th-edition
[B261] Wabner D, Beier C. Aromaterapi: grunder, verkningsmekanismer, praktik. 2:a uppl. Urban & Fischer; 2012.https://www.elsevier.com/books/aromatherapie/wabner/978-3-437-57214-4
[B262] Lis-Balchin M. Geranium och Pelargonium: Släktena Geranium och Pelargonium. CRC Press; 2002.https://www.routledge.com/Geranium-and-Pelargonium/Lis-Balchin/p/book/9780415284653
[B263] Turek C, Stintzing FC. Stabilitet hos eteriska oljor: En översikt. Compr Rev Food Sci Food Saf. 2013;12(1):40-53.https://doi.org/10.1111/1541-4337.12006
[B264] Skold M, Karlberg AT, Matura M, Börje A. Doftkemikalien beta-karyofyllen – luftoxidation och hudsensibilisering. Food Chem Toxicol. 2006;44(4):538-545.https://doi.org/10.1016/j.fct.2005.08.028
[B265] Tavassoli T, Miller LJ, Schoen SA, Nielsen DM, Baron-Cohen S. Sensorisk överkänslighet hos vuxna med autismspektrumtillstånd. Autism. 2014;18(4):428-432.https://doi.org/10.1177/1362361313477246
[B266] Lane AE, Young RL, Baker AE, Angley MT. Sensory processing subtypes in autism: association with adaptive behavior. J Autism Dev Disord. 2010;40(1):112-122.https://doi.org/10.1007/s10803-009-0840-2
[B267] Mazurek MO, Vasa RA, Kalb LG, et al. Ångest, sensorisk överkänslighet och mag-tarmproblem hos barn med autism-spektrumstörningar. J Abnorm Child Psychol. 2013;41(1):165-176.https://doi.org/10.1007/s10802-012-9668-x
[B268] Buckle J. Klinisk aromaterapi: eteriska oljor i praktiken. 2:a uppl. Churchill Livingstone; 2003.https://www.elsevier.com/books/clinical-aromatherapy/buckle/978-0-7020-2536-6
[B269] Tuchman R, Cuccaro M, Alessandri M. Autism och epilepsi: historiskt perspektiv. Brain Dev. 2010;32(9):709-718.https://doi.org/10.1016/j.braindev.2010.04.008
[B270] Burkhard PR, Burkhardt K, Haenggeli CA, Landis T. Krapinducerade anfall: återkomst av ett gammalt problem. J Neurol. 1999;246(8):667-670.https://doi.org/10.1007/s004150050434
[B271] Millet Y, Jouglard J, Steinmetz MD, Tognetti P, Joanny P, Arditti J. Toxicitet hos några eteriska oljor från växter. Klinisk och experimentell studie. Clin Toxicol. 1981;18(12):1485-1498.https://doi.org/10.3109/15563658108990111
[B272] Pengelly A, Bennett K, Spinks J, Bensoussan A. Appraising the evidence-base for the use of essential oils in the management of epilepsy: A systematic review. Epilepsy Behav. 2012;24(1):9-15.https://doi.org/10.1016/j.yebeh.2012.02.019
Höld KM, Sirisoma NS, Ikeda T, Narahashi T, Casida JE. Alfa-tujon (den aktiva komponenten i absint): modulering av GABA A-receptorn och metabolisk avgiftning. Proc Natl Acad Sci U S A. 2000;97(8):3826-3831.https://doi.org/10.1073/pnas.070042397
[B274] Hyssopus-olja och salviaolja – studier av krampframkallande egenskaper hos kommersiella prover av eteriska oljor från Hyssopus officinalis L. och Salvia officinalis L. Plantes Med Phytother. 1979;13(3):179-184.https://www.sciencedirect.com/science/article/abs/pii/S0370447579800477
[B275] Marcus C, Lichtenstein EP. Interaktioner mellan naturligt förekommande komponenter i matväxter och insektsmedel samt pentobarbital hos råttor och möss. J Agric Food Chem. 1979;27(5):1217-1223.https://doi.org/10.1021/jf60226a004
[B276] Koutroumanidou E, Kimbaris A, Kortsaris A, et al. Ökad anfallslatens och minskad svårighetsgrad av pentylentetrazol-inducerade anfall hos möss efter administrering av eterisk olja. Epilepsy Res Treat. 2013;2013:532657.https://doi.org/10.1155/2013/532657
[B277] Avallone R, Zanoli P, Puia G, Kleinschnitz M, Schreier P, Baraldi M. Farmakologiska profil av apigenin, en flavonoid isolerad från Matricaria chamomilla. Biochem Pharmacol. 2000;59(11):1387-1394.https://doi.org/10.1016/s0006-2952(00 )00264-1
[B278] Watanabe E, Kuchta K, Kimura M, Rauwald HW, Kamei T, Imanishi J. Effekter av eterisk olja från bergamott (Citrus bergamia (Risso) Wright & Arn.) inom aromaterapi på sinnesstämningar, parasympatiska nervsystemets aktivitet och salivkortisolnivåer hos 41 friska kvinnor. Forsch Komplementmed. 2015;22(1):43-49.https://doi.org/10.1159/000380989
[B279] Ilhan S, Savaşer S, Kölgelier S. Aromaterapins effekt på patienter med epilepsi. Epilepsy Behav. 2004;5(5):753-757.https://doi.org/10.1016/j.yebeh.2004.06.004
[B280] Meamarbashi A, Rajabi A. Pepparmyntans effekter på träningsprestation. J Int Soc Sports Nutr. 2013;10(1):15.https://doi.org/10.1186/1550-2783-10-15
[B281] Eccles R. Mentol och relaterade kylpreparat. J Pharm Pharmacol. 1994;46(8):618-630.https://doi.org/10.1111/j.2042-7158.1994.tb03871.x
[B282] Silva J, Abebe W, Sousa SM, Duarte VG, Machado MI, Matos FJ. Analgetiska och antiinflammatoriska effekter av eteriska oljor från eukalyptus. J Ethnopharmacol. 2003;89(2-3):277-283.https://doi.org/10.1016/j.jep.2003.09.007
[B283] Fachini-Queiroz FC, Kummer R, Estevão-Silva CF, et al. Effekter av tymol och karvakrol, beståndsdelar i eterisk olja från Thymus vulgaris L., på inflammatoriska svar. Evid Based Complement Alternat Med. 2012;2012:657026.https://doi.org/10.1155/2012/657026
[B284] Carson CF, Hammer KA, Riley TV. Melaleuca alternifolia (Tea Tree) Oil: a Review of Antimicrobial and Other Medicinal Properties. Clin Microbiol Rev. 2006;19(1):50-62.https://doi.org/10.1128/CMR.19.1.50-62.2006
[B285] Eccles R, Jawad MS, Morris S. Effekterna av oral administrering av (-)-mentol på näsans luftmotstånd och näsans känsla av luftflöde hos försökspersoner som lider av nästäppa förknippad med vanlig förkylning. J Pharm Pharmacol. 1990;42(9):652-654.https://doi.org/10.1111/j.2042-7158.1990.tb06625.x
[B286] Altunbas MK, Celık F, Kahraman C. Lavendeloljans potential för säker användning hos nyfödda. J Neonatal Nurs. 2020;26(6):303-307.https://doi.org/10.1016/j.jnn.2020.05.005
[B287] Srivastava JK, Shankar E, Gupta S. Kamomill: Ett växtbaserat läkemedel från det förflutna med ljus framtid. Mol Med Rep. 2010;3(6):895-901.https://doi.org/10.3892/mmr.2010.377
[B288] Kiecolt-Glaser JK, Graham JE, Malarkey WB, Porter K, Lemeshow S, Glaser R. Luktets inverkan på humör och autonom, endokrin och immun funktion. Psychoneuroendocrinology. 2008;33(3):328-339.https://doi.org/10.1016/j.psyneuen.2007.11.015
[B289] Halm MA. Eteriska oljor för hantering av symtom hos allvarligt sjuka patienter. Am J Crit Care. 2008;17(2):160-163.https://www.researchgate.net/publication/5541002_Essential_Oils_for_Management_of_Symptoms_in_Critically_Ill_Patients
[B290] Fung LK, Mahajan R, Nozzolillo A, et al. Farmakologisk behandling av svår irritabilitet och problembeteenden vid autism: En systematisk översikt och metaanalys. Pediatrics. 2016;137 Suppl 2:S124-S135.https://doi.org/10.1542/peds.2015-2851K
[B291] Kirschbaum KM, Müller MJ, Malevani J, et al. Serumkoncentrationer av aripiprazol och dehydroaripiprazol, kliniskt svar och biverkningar. World J Biol Psychiatry. 2008;9(3):212-218.https://doi.org/10.1080/15622970701361255
[B292] Sprouse J, Clarke T, Reynolds L, Heym J, Rollema H. Jämförelse av effekterna av sertralin och dess metabolit desmetylsertralin på blockad av central 5-HT-återupptag in vivo. Neuropsychopharmacology. 1996;14(4):225-231.https://doi.org/10.1016/0893-133X(95 )00112-Q
[B293] Bailey DG, Dresser G, Arnold JM. Grapefrukt-läkemedelsinteraktioner: förbjuden frukt eller undvikbara konsekvenser? CMAJ. 2013;185(4):309-316.https://doi.org/10.1503/cmaj.120951
[B294] Obach RS, Walsky RL, Venkatakrishnan K, Gaman EA, Houston JB, Tremaine LM. The utility of in vitro cytochrome P450 inhibition data in the prediction of drug-drug interactions. J Pharmacol Exp Ther. 2006;316(1):336-348.https://doi.org/10.1124/jpet.105.093229
[B295] Guo LQ, Yamazoe Y. Inhibition av cytokrom P450 av furanokumariner i grapefruktjuice och örtmediciner. Acta Pharmacol Sin. 2004;25(2):129-136.https://www.researchgate.net/publication/8881220_Inhibition_of_cytochrome_P450_by_furanocoumarins_in_grapefruit_juice_and_herbal_medicines
[B296] Paine MF, Criss AB, Watkins PB. Två huvudkomponenter i grapefruktjuice skiljer sig i tid till debut av intestinal CYP3A4-hämning. J Pharmacol Exp Ther. 2004;312(3):1151-1160.https://doi.org/10.1124/jpet.104.076570
[B297] Hanley MJ, Cancalon P, Widmer WW, Greenblatt DJ. Effekten av grapefruktjuice på läkemedelsmetabolism. Expert Opin Drug Metab Toxicol. 2011;7(3):267-286.https://doi.org/10.1517/17425255.2011.553189
[B298] Dresser GK, Bailey DG. Effekterna av fruktjuicer på läkemedels disposition: en ny modell för läkemedelsinteraktioner. Eur J Clin Invest. 2003;33 Suppl 2:10-16.https://doi.org/10.1046/j.1365-2362.33.s2.2.x
[B299] Izzo AA, Ernst E. Interaktioner mellan örtmediciner och föreskrivna läkemedel: en uppdaterad systematisk översikt. Drugs. 2009;69(13):1777-1798.https://doi.org/10.2165/11317010-000000000-00000
[B300] Henderson L, Yue QY, Bergquist C, Gerden B, Arlett P. Johannesört (Hypericum perforatum): läkemedelsinteraktioner och kliniska resultat. Br J Clin Pharmacol. 2002;54(4):349-356.https://doi.org/10.1046/j.1365-2125.2002.01683.x
[B301] Unger M, Frank A. Samtidig bestämning av den hämmande potens av örtsextrakt på aktiviteten av sex stora cytokrom P450-enzymer med hjälp av vätskekromatografi/masspektrometri och automatiserad online-extraktion. Rapid Commun Mass Spectrom. 2004;18(19):2273-2281.https://doi.org/10.1002/rcm.1621
[B302] Gurley BJ, Gardner SF, Hubbard MA, et al. In vivo-effekter av goldenseal, kava kava, svart cohosh och valeriana på mänskliga cytokrom P450 1A2, 2D6, 2E1 och 3A4/5 fenotyper. Clin Pharmacol Ther. 2005;77(5):415-426.https://doi.org/10.1016/j.clpt.2005.01.009
[B303] Spinella M. Betydelsen av farmakologisk synergi i psykoaktiva växtbaserade läkemedel. Altern Med Rev. 2002;7(2):130-137.https://pubmed.ncbi.nlm.nih.gov/11968843/
[B304] Abourashed EA, Koetter U, Brattström A. In vitro-bindningsexperiment med en extrakt av valeriana, humle och deras fasta kombination (Ze91019) till utvalda centrala nervsystemets receptorer. Phytomedicine. 2004;11(7-8):633-638.https://doi.org/10.1016/j.phymed.2004.03.005
[B305] Brazier NC, Levine MA. Läkemedelsinteraktioner mellan vanliga konventionella läkemedel: en kompendium för hälso- och sjukvårdspersonal. Am J Ther. 2003;10(3):163-169.https://doi.org/10.1097/00045391-200305000-00003
[B306] De Groot AC, Schmidt E. Essential oils: contact allergy and chemical composition. CRC Press; 2016.https://www.routledge.com/Essential-Oils-Contact-Allergy-and-Chemical-Composition/de-Groot-Schmidt/p/book/9781498702508
[B307] Shellie R, Marriott P, Huie C. Comprehensive tvådimensionell gaskromatografi (GC × GC) och GC × GC-kvadrupol-MS-analys av asiatisk och amerikansk ginseng. J Sep Sci. 2003;26(12-13):1185-1192.https://doi.org/10.1002/jssc.200301521
[B308] Kubeczka KH. History and sources of essential oil research. I: Baser KHC, Buchbauer G, red. Handbook of Essential Oils: Science, Technology, and Applications. CRC Press; 2010:3-38.https://www.routledge.com/Handbook-of-Essential-Oils-Science-Technology-and-Applications/Baser-Buchbauer/p/book/9781032046419
[B309] Cavanagh HM, Wilkinson JM. Lavendel eterisk olja: en översikt. Aust Infect Control. 2005;10(1):35-37.https://doi.org/10.1071/HI05035
[B310] Leung AY, Foster S. Encyclopedia of Common Natural Ingredients Used in Food, Drugs, and Cosmetics. 2nd ed. Wiley; 1996.https://www.wiley.com/en-us/Encyclopedia+of+Common+Natural+Ingredients+Used+in+Food%2C+Drugs%2C+and+Cosmetics%2C+2nd+Edition-p-9780471508267
[B311] Lis-Balchin M. Aromaterapi med eteriska oljor. I: Lis-Balchin M, red. Aromatherapy Science: A Guide for Healthcare Professionals. Pharmaceutical Press; 2006:49-115.https://www.pharmpress.com/product/9780853696230/aromatherapy-science
[B312] Williams TI. En orolig blick på aromaterapi. Perspect Psychiatr Care. 2006;42(2):107-108.https://doi.org/10.1111/j.1744-6163.2006.00056.x
[B313] Brondino N, Fusar-Poli L, Rocchetti M, Provenzani U, Barale F, Politi P. Komplementära och alternativa terapier för autismspektrumstörning. Evid Based Complement Alternat Med. 2015;2015:258589.https://doi.org/10.1155/2015/258589
[B314] Bent S, Bertoglio K, Hendren RL. Omega-3-fettsyror för autism för att förstå spektrumet: en systematisk översikt. J Autism Dev Disord. 2009;39(8):1145-154.https://doi.org/10.1007/s10803-009-0724-5
[B315] Masi A, Quintana DS, Glozier N, Lloyd AR, Hickie IB, Guastella AJ. Cytokineavvikelser vid autism-spektrumstörning: en systematisk översikt och metaanalys. Mol Psychiatry. 2015;20(4):440-446.https://doi.org/10.1038/mp.2014.59
[B316] Veenstra-VanderWeele J, Blakely RD. Networking in autism: leveraging genetic, biomarker and model system findings in the search for new treatments. Neuropsychopharmacology. 2012;37(1):196-212.https://doi.org/10.1038/npp.2011.185
[B317] Yamasue H, Domes G. Oxytocin och autismspektrumstörningar. Curr Top Behav Neurosci. 2018;35:449-465.https://doi.org/10.1007/7854_2017_24
[B318] Petra AI, Panagiotidou S, Hatziagelaki E, Stewart JM, Conti P, Theoharides TC. Tarm-Mikrobiota-Hjärna-Axeln och dess effekt på neuropsychiatriska sjukdomar med misstänkt immun-dysreglering. Clin Ther. 2015;37(5):984-995.https://doi.org/10.1016/j.clinthera.2015.04.002
[B319] Mayer EA, Tillisch K, Gupta A. Tarm/hjärna-axeln och mikrobiotan. J Clin Invest. 2015;125(3):926-938.https://doi.org/10.1172/JCI76304
[B320] Button KS, Ioannidis JP, Mokrysz C, et al. Power failure: why small sample size undermines the reliability of neuroscience. Nat Rev Neurosci. 2013;14(5):365-376.https://doi.org/10.1038/nrn3475
[B321] Higgins JP, Thompson SG, Deeks JJ, Altman DG. Mätning av inkonsekvens i metaanalyser. BMJ. 2003;327(7414):557-560.https://doi.org/10.1136/bmj.327.7414.557
[B322] Rossignol DA. Nya och framväxande behandlingar för autism-spektrumstörningar: en systematisk översikt. Ann Clin Psychiatry. 2009;21(4):213-236.https://pmc.ncbi.nlm.nih.gov/articles/PMC3859980/
[B323] Moher D, Liberati A, Tetzlaff J, Altman DG; PRISMA Group. Preferred reporting items for systematic reviews and meta-analyses: the PRISMA statement. PLoS Med. 2009;6(7):e1000097.https://doi.org/10.1371/journal.pmed.1000097
[B324] Ioannidis JP, Patsopoulos NA, Rothstein HR. Anledningar eller ursäkter för att undvika metaanalys i skogsplottar. BMJ. 2008;336(7658):1413-1415.https://doi.org/10.1136/bmj.a117
[B325] Loth E, Charman T, Mason L, et al. EU-AIMS Longitudinal European Autism Project (LEAP): design och metoder för att identifiera och validera stratifieringsbiomarkörer för autism-spektrumstörningar. Mol Autism. 2017;8:24.https://doi.org/10.1186/s13229-017-0146-8
[B326] Geschwind DH, Flint J. Genetics and genomics of psychiatric disease. Science. 2015;349(6255):1489-1494.https://doi.org/10.1126/science.aaa8954
[B327] Easterbrook PJ, Berlin JA, Gopalan R, Matthews DR. Publiceringsbias inom klinisk forskning. Lancet. 1991;337(8746):867-872.https://doi.org/10.1016/0140-6736(91 )90201-y
[B328] Nestler EJ, Hyman SE. Djurmodeller för neuropsykiatriska sjukdomar. Nat Neurosci. 2010;13(10):1161-1169.https://doi.org/10.1038/nn.2647
[B329] Buchbauer G, Jirovetz L, Jäger W, Plank C, Dietrich H. Doftämnen och eteriska oljor med lugnande effekt vid inandning. J Pharm Sci. 1993;82(6):660-664.https://doi.org/10.1002/jps.2600820623
[B330] Cooke B, Ernst E. Aromaterapi: en systematisk översikt. Br J Gen Pract. 2000;50(455):493-496.https://pubmed.ncbi.nlm.nih.gov/10962794/
[B331] Halm MA. Eteriska oljor för hantering av symtom hos kritiskt sjuka patienter. Am J Crit Care. 2008;17(2):160-163.https://www.researchgate.net/publication/5541002_Essential_Oils_for_Management_of_Symptoms_in_Critically_Ill_Patients
[B332] Hammer KA, Carson CF, Riley TV. Antimikrobiell aktivitet av eteriska oljor och andra växtextrakt. J Appl Microbiol. 1999;86(6):985-990.https://doi.org/10.1046/j.1365-2672.1999.00780.x
[B333] Schulz KF, Altman DG, Moher D; CONSORT Group. CONSORT 2010 statement: uppdaterade riktlinjer för rapportering av randomiserade studier med parallella grupper. BMJ. 2010;340:c332.https://doi.org/10.1136/bmj.c332
[B334] Lombardo MV, Lai MC, Baron-Cohen S. Big data approaches to decomposing heterogeneity across the autism spectrum. Mol Psychiatry. 2019;24(10):1435-1450.https://doi.org/10.1038/s41380-018-0321-0
[B335] Insel TR. NIMH Research Domain Criteria (RDoC) Projektet: precisionsmedicin för psykiatrin. Am J Psychiatry. 2014;171(4):395-397.https://doi.org/10.1176/appi.ajp.2014.14020138
[B336] Jeste SS, Geschwind DH. Kliniska prövningar för neurodevelopmentala störningar: På en terapeutisk frontlinje. Sci Transl Med. 2016;8(321):321fs1.https://doi.org/10.1126/scitranslmed.aad9874
[B337] Kovar KA, Gropper B, Friess D, Ammon HP. Blodnivåer av 1,8-cineol och lokomotorisk aktivitet hos möss efter inhalation och oral administrering av rosmarinolja. Planta Med. 1987;53(4):315-318.https://doi.org/10.1055/s-2006-962725
[B338] Rossignol DA, Frye RE. En översikt av forskningstrender inom fysiologiska abnormaliteter vid autismspektrumstörningar: immunregleringsstörningar, inflammation, oxidativ stress, mitokondriell dysfunktion och exponering för miljögifter. Mol Psychiatry. 2012;17(4):389-401.https://doi.org/10.1038/mp.2011.165
[B339] Stang A. Kritisk utvärdering av Newcastle-Ottawa-skalan för bedömning av kvaliteten på icke-randomiserade studier i metaanalyser. Eur J Epidemiol. 2010;25(9):603-605.https://doi.org/10.1007/s10654-010-9491-z
[B340] Herz RS. Aromaterapifakta och fiktion: en vetenskaplig analys av lukteffekter på humör, fysiologi och beteende. Int J Neurosci. 2009;119(2):263-290.https://doi.org/10.1080/00207450802333953
[B341] Myers SM, Johnson CP; American Academy of Pediatrics Council on Children With Disabilities. Hantering av barn med autismspektrumstörningar. Pediatrics. 2007;120(5):1162-1182.https://doi.org/10.1542/peds.2007-2362
[B342] Volkmar F, Siegel M, Woodbury-Smith M, King B, McCracken J, State M; American Academy of Child and Adolescent Psychiatry (AACAP) Committee on Quality Issues (CQI). Practice parameter for the assessment and treatment of children and adolescents with autism spectrum disorder. J Am Acad Child Adolesc Psychiatry. 2014;53(2):237-257.https://doi.org/10.1016/j.jaac.2013.10.013
[B343] Green VA, Pituch KA, Itchon J, Choi A, O’Reilly M, Sigafoos J. Internetenkät om behandlingar som används av föräldrar till barn med autism. Res Dev Disabil. 2006;27(1):70-84.https://doi.org/10.1016/j.ridd.2004.12.002
[B344] Bilia AR, Guccione C, Isacchi B, Righeschi C, Firenzuoli F, Bergonzi MC. Eteriska oljor inneslutna i nanosystem: en utvecklingsstrategi för ett framgångsrikt terapeutiskt tillvägagångssätt. Evid Based Complement Alternat Med. 2014;2014:651593.https://doi.org/10.1155/2014/651593
[B345] Woolf AD. Essentiell oljeförgiftning. J Toxicol Clin Toxicol. 1999;37(6):721-727.https://doi.org/10.1081/clt-100102450
[B346] Kasper S, Gastpar M, Müller WE, et al. Effektivitet och säkerhet av Silexan, en ny, oralt administrerad lavendeloljepreparat, vid subtröskel ångeststörning – bevis från kliniska prövningar. Wien Med Wochenschr. 2010;160(21-22):547-556.https://doi.org/10.1007/s10354-010-0845-7
B347. Ingefärstillskott lindrar autistiskt beteende genom att modulera AKT/GSK3β-signalering hos möss som exponerats för prenatalt valproinsyra. PubMed. https://doi.org/10.1039/d5fo04364f
B348. Effekt av Hiwa-sirap, en persisk medicinprodukt, på autismssymtom hos barn med autismspektrumstörningar: En randomiserad dubbelblindad klinisk prövning. Global Medicinsk Tidskrift. https://doi.org/10.31661/gmj.v13i.3553
B349. McGuinness, F., Oswald, I. W. H., Koby, K. A., m.fl. (2023). Cannabidiol och cannabis-inspirerade terpenblandningar har akuta prosociala effekter i BTBR-musmodellen av autism. Frontiers in Neuroscience. https://doi.org/10.3389/fnins.2023.1185737
ADHD Ordbok
ADHD (Attention Deficit Hyperactivity Disorder)
Neuropsykiatrisk sjukdom, kännetecknad av ouppmärksamhet, hyperaktivitet och impulsivitet.
Adjuverande behandling
Tilläggsbehandling som används utöver huvudbehandlingen för att förstärka dess effekt eller minska biverkningar.
Ångestdämpande
Ångestdämpande, ångestrelieferande.
Blod-hjärnbarriären
Selektiv barriär mellan blodbanan och hjärnan, som kontrollerar passage av substanser in i hjärnan.
CB2-receptorn
Kannabinoidreceptor typ 2, en del av endocannabinoidsystemet, finns primärt på immunceller; aktivering har antiinflammatorisk verkan.
Kemotyp
Kemisk variant av en växt med olika sammansättning av eteriska oljor (t.ex. rosmarin kemotyp cineol vs. kamfer).
DAT (dopamintransportör)
Protein som transporterar dopamin från den synaptiska klyftan tillbaka till nervcellen; huvudmål för metylfenidat.
DAT-ockupanz
Andel dopamintransportproteiner som blockeras av ett läkemedel.
Dopamin
Neurotransmittor, viktig för motivation, belöning, rörelse och uppmärksamhet; ofta nedsatt vid ADHD.
ED₅₀
Dos, vid vilken 50% av maximal effekt uppnås.
GABA (gamma-aminosmörsyra)
Viktigaste hämmande signalsubstansen i hjärnan; minskar neuronal retbarhet.
GABA-A-receptor
GABA-jonkanalkanalreceptor; aktivering leder till kloridinflöde och hyperpolarisering (hämning) av nervcellen.
GC/MS (Gaskromatografi-masspektrometri)
Analytisk metod för identifiering och kvantifiering av kemiska komponenter i eteriska oljor.
IC₅₀
Koncentration vid vilken 50% av maximal hämning av ett mål (t.ex. receptor, enzym) uppnås.
Limbiska systemet
Hjärnregion, som bearbetar känslor, minne och motivation; omfattar amygdala, hippocampus och andra strukturer.
Linalool
Monoterpenalkohol, huvudsaklig aktiv ingrediens i lavendel; har ångestdämpande, lugnande effekter och modulerar signalsubstanssystem.
Mentol
Monoterpenalkohol från pepparmynta; aktiverar TRPM8-köldreceptorer, verkar uppfriskande och uppmärksamhetsstimulerande.
Metylfenidat (MPH)
Stimulans, som hämmar återupptaget av dopamin och noradrenalin; förstahandsbehandling vid ADHD.
Mikrodialys
Teknik för att mäta extracellulära neurotransmittorkoncentrationer i levande hjärna.
Monoamin
Grupp av signalsubstanser som inkluderar dopamin, noradrenalin och serotonin.
NET (Noradrenalintransportör)
Protein som transporterar noradrenalin från den synaptiska klyftan tillbaka till nervcellen; även måltavla för metylfenidat.
Neuroinflammation
Inflammatoriska processer i hjärnan, ofta medierade av aktiverade mikroglia och proinflammatoriska cytokiner.
Neurotransmittorer
Kemiska budbärare som överför signaler mellan nervceller (t.ex. dopamin, serotonin, GABA).
NF-κB (kärnfaktor kappa B)
Transkriptionsfaktor som aktiverar inflammatoriska gener; hämning verkar antiinflammatoriskt.
NMDA-receptorn
Glutamat-receptor, viktig för synaptisk plasticitet och inlärning; överaktivering kan vara neurotoxiskt.
Noradrenalin
Neurotransmittor, viktig för vakenhet, uppmärksamhet och stressrespons; ofta nedsatt vid ADHD.
Ockupanz
Se DAT-ockupans; procentandel belagda receptorer eller transportörer.
Luktsinnet
Vad gäller luktsinnet.
PET (Positronemissionstomografi)
Bildgivande metod som använder radioaktivt märkta substanser för att visualisera metaboliska processer och receptorbindning i hjärnan.
Farmakokinetik
Lär dig vad kroppen gör med ett läkemedel (absorption, distribution, metabolism, utsöndring).
Farmakodynamik
Lär dig om hur ett läkemedel verkar i kroppen (verkningsmekanismer, effekter).
Prefrontala cortex (PFC)
Hjärnregion bakom pannan, viktig för exekutiva funktioner (planering, impulskontroll, arbetsminne); ofta underaktiv vid ADHD.
Receptor
Proteiner på eller i celler, som binder specifika signalmolekyler (neurotransmittorer, hormoner) och därmed utlöser cellulära reaktioner.
SERT (Serotonin-transportör)
Protein som transporterar serotonin från den synaptiska klyftan tillbaka in i nervcellen; mål för antidepressiva medel (SSRI).
Serotonin
Neurotransmittor, viktig för humör, sömn, aptit och impulskontroll.
SPECT (Single Photon Emission Computed Tomography
Bildgivande metod liknande PET, som använder radioaktiva spårämnen för att avbilda hjärnfunktioner.
Striatum
Hjärnregion i basala ganglierna, rik på dopamintransportörer; viktig för rörelse, motivation och belöning; huvudmål för metylfenidat.
Synaps
Synaps.
Synapsspalt
Smalt utrymme mellan två nervceller vid synapsen, i vilket signalsubstanser frisätts.
TRP-kanaler (Transient Receptor Potential)
Familj av jonkanaler som reagerar på olika stimuli (temperatur, kemikalier); t.ex. TRPM8 (kyla/mentol), TRPV1 (värme/kapsaicin).
Venturiprincipen
Fysikaliskt princip där vätskor eller oljor sugs in och finfördelas genom tryckfall i en förträngning; grunden för många diffusorer.
β-karyofyllen
Sesquiterpener från svartpeppar och cannabis; selektiv CB2-receptoragonist med antiinflammatoriska egenskaper.
1,8-Cineol (Eukalyptol)
Monoterpenoxid från rosmarin och eukalyptus; verkar slemlösande, antiinflammatoriskt och kognitivt stimulerande.
5-HT1A-receptor
Serotoninreceptorsubtyp; aktivering verkar ångestdämpande och antidepressiv.
Rapportens slut
Denna rapport skapades den 27 april 2026 baserat på en systematisk analys av 87 studier om metylfenidat-hjärnkoncentrationer och 159 publikationer om eteriska oljor i ADHD-sammanhang. Alla uttalanden stöds av primärlitteratur.
Ordlista ASD
1,8-Cineol (Eukalyptol)
Monoterpenoxid, huvudkomponent i eukalyptusolja (70–90 %), uppvisar ångestdämpande effekter via GABA-A-receptormodulering och antiinflammatoriska egenskaper; kontraindicerat för barn under 3 år på grund av risk för struplockskramper.
α-Pinen
Bicyklisk monoterpen i tall, rosmarin och cannabis; visar GABAerg modulering, ångestdämpande och antiinflammatoriska effekter; preklinisk evidens för neuroprotektiva egenskaper.
Amygdala
Mandelformad hjärnstruktur i tinningloben, central för känslomässig bearbetning, rädsloinlärning och social perception; förstorad i tidig barndom vid ASD, minskad aktivering vid ansiktsbearbetning.
Antioxidant
Molekyl som minskar oxidativ stress genom att neutralisera reaktiva syreföreningar (ROS); exempel: glutation, superoxiddismutas, katalas, vitamin C, vitamin E; minskad antioxidativ kapacitet vid ASD.
Tillämpad beteendeanalys (ABA)
Evidensbaserad beteendeterapeutisk intervention för ASD, baserad på inlärningsprinciper (förstärkning, prompting, shaping); förbättrar kommunikation, sociala färdigheter och adaptiva beteenden.
Aripiprazol
Atypiskt antipsykotikum, partiell dopamin-D2-agonist; FDA-godkänt för irritabilitet vid ASD; Biverkningar: viktökning, akatisi, metaboliskt syndrom; metaboliseras via CYP2D6 och CYP3A4.
Astrocyter
Stjärnformiga gliaceller i CNS; reglerar glutomhäradhet (via EAAT1/2-transportörer), K+-buffring, blod-hjärnbarriären och metabolt stöd till neuroner; vid ASD astroglios (ökad GFAP-expression).
β-karyofyllen (BCP)
Sesquiterpenkolväte i svartpeppar, kryddnejlika, cannabis; selektiv CB2-receptoragonist utan psykoaktiva effekter; minskar neuroinflammation, mikrogliaaktivering och oxidativ stress.
Bensodiazepinbindningsställe
Allosterisk modulationsplats på GABA-A-receptorn; bensodiazepiner (diazepam, lorazepam) binder här och förstärker GABAerg hämning; linalool och 1,8-cineol interagerar med denna plats (flumazenil-känslig).
Bergamott (Citrus bergamia)
Citrusfrukt, eterisk olja innehåller linalool, linalylacetat, limonen; en RCT på barn med ASD visade inga signifikanta ångestdämpande effekter; innehåller bergapten (CYP3A4-hämmare), bergaptenfria varianter (FCF) tillgängliga.
Boswelliasyror
Pentacykliska triterpensyror från Boswellia (rökelse); aktiverar Nrf2/HO-1-antioxidantvägen, minskar neuroinflammation (mikroglia, astrocyter, TNF-α, IL-6); prekliniska bevis för förbättring av ASD-liknande beteenden i VPA-modeller.
Bumetanid
Loopdiuretikum, NKCC1-kotransportörhämmare; korrigerar fördröjd GABA-växling i ASD-mousmodeller; kliniska studier visar måttliga förbättringar i social kommunikation hos ASD-barn, men med metodologiska begränsningar.
Kamfer
Terpenketon i kamfer (Cinnamomum camphora) och rosmarin (kamfer-kemotyp); sänker kramptröskeln, kan utlösa anfall; kontraindicerat vid epilepsi och patienter med ASD med komorbid epilepsi.
Cananga odorata (Ylang-Ylang)
Tropiskt träd, eterisk olja innehåller linalool, geraniol, β-karyofyllen; preklinisk studie visade förbättring av ASD-liknande beteenden hos VPA-ratta genom modulering av serotonin- och dopaminmetabolism.
CB2-receptorn (Cannabinoid-typ-2-receptor)
G-proteinkopplad receptor, primärt uttryckt på immunceller (mikroglia, perifera immunceller); aktivering minskar cytokinfrisättning, mikrogliaaktivering och neuroinflammation; β-karyofyllen är en selektiv CB2-agonist.
Cedrol
Sesquiterpenalkohol i cederolje; traditionellt associerad med lugnande egenskaper, men vetenskapliga bevis för GABAerga mekanismer eller klinisk effekt vid ASD saknas.
Kopienumre varianter (CNV)
Genomiska deletioner eller duplikationer >1 kb; vanliga vid ASD (15q11-13-duplikation, 16p11.2-deletion, 22q13-deletion/Phelan-McDermid syndrom); involverar ofta gener relaterade till synapser (SHANK3, NLGN4).
Cytochrom P450 (CYP)
Enzymfamilj som katalyserar läkemedelsmetabolism; CYP3A4 och CYP2D6 metaboliserar risperidon och aripiprazol; bergapten (i bergamott) hämmar CYP3A4, kan öka läkemedelsnivåer.
Dopamin (DA)
Katekolamin-neurotransmittor; förmedlar belöningsbearbetning, motivation, motorisk kontroll; vid ASD reducerad striatal aktivering vid sociala belöningar, förändrad dopamintransportdensitet; Ylang-Ylang modulerar DA-metabolism i prekliniska modeller.
DSM-5 (Diagnostic and Statistical Manual of Mental Disorders, 5:e upplagan)
Klassifikationssystem från American Psychiatric Association; definierar ASD genom två huvuddomäner: (1) bestående brister i social kommunikation/interaktion, (2) begränsade, repetitiva beteendemönster.
Dysbios
Obalans i tarmmikrobiotan; vid ASD reducerad mikrobiell diversitet, ökat Firmicutes/Bacteroidetes-förhållande, reducerat Bifidobacterium och Prevotella; korrelerar med symtomens svårighetsgrad.
E/I-obalans (Excitatorisk-Inhibitorisk obalans)
Obalans mellan excitatorisk (glutamaterg) och inhibitorisk (GABAerg) neurotransmission; central patofysiologisk hypotes vid ASD; leder till hyperexcitabilitet, störd sensorisk filtrering och nedsatt social informationsbehandling.
Elektroakupunktur
Akupunktur med elektrisk stimulering; prekliniska studier visar förbättring av ASD-liknande beteenden hos VPA-möss via vagusberoende mekanismer och mikrobiommodulering.
Epilepsi
Neurologische Erkrankung mit rekurrenten Anfällen; 10–30 % der ASD-Patienten haben komorbide Epilepsie; campherhaltige ätherische Öle (Kampfer, Rosmarin-Campher-Chemotyp, Salbei) sind kontraindiziert.
Fekal mikrobiotatransplantation (FMT)
Överföring av tarmmikrobiota från friska donatorer till mottagare; prekliniska studier visade förbättring av ASD-liknande beteenden i musmodeller; öppen klinisk studie på ASD-barn visade förbättringar i GI-symtom och beteende.
Flumazenil
Bensodiazepinantagonist, binder till bensodiazepinbindningsstället på GABA-A-receptorn; blockerar anxiolytiska effekter av linalool och 1,8-cineol, vilket bekräftar deras interaktion med denna plats.
GABA (gamma-aminosmörsyra)
Primära hämmande signalsubstansen i CNS; binder till GABA-A- (jonotropha) och GABA-B-receptorer (metabotropa); vid ASD reducerad GABAerg funktion, förändrade GABA-nivåer, störd GABA-switch under utveckling.
GABA-A-receptor
Ligandstyrd kloridkanal; aktivering av GABA leder till inströmning av kloridjoner och hyperpolarisering (hämning); innehåller bensodiazepinbindningsställe; linalool, 1,8-cineol och α-pinen modulerar GABA-A-receptorer.
GABA-omkopplare
Utvecklingsbiologisk övergång från excitatorisk till inhiberande GABA-effekt; medieras av en förändring i uttrycket av NKCC1- till KCC2-kotransporter; försenad vid ASD; oxytocin och bumetanid kan normalisera GABA-omkopplingen.
GAD65/GAD67 (Glutamat-dekarboxylas 65/67)
Enzymer som omvandlar glutamat till GABA; vid ASD reducerad GAD65/67-expression i prefrontala cortex och cerebellum, vilket leder till reducerad GABA-syntes.
Gaskromatografi-masspektrometri (GC/MS)
Analytisk teknik för identifiering och kvantifiering av flyktiga komponenter; guldstandard för kemisk karakterisering av eteriska oljor; möjliggör detektering av förfalskningar, föroreningar och variationer mellan batcher.
Glutamat (Glu)
Primär excitatorisk neurotransmittor i CNS; binder till jonotropa (NMDA, AMPA, Kainat) och metabotropa (mGluR1-8) receptorer; vid ASD förändrade glutamatnivåer, störda glutamaterga synapse-gener (GRIN2B, GRIA1), överdriven glutamaterg transmission.
Glutation (GSH)
Tripeptid (γ-Glutamyl-Cysteinyyl-Glycin), primärt intracellulärt antioxidant; vid ASD reducerade GSH-nivåer, ökat oxiderat glutation (GSSG), reducerat GSH/GSSG-förhållande; genetiska varianter i GSH-syntesenzymer (GCLC, GSS) associerade med ASD.
Tarm-hjärna-axeln
Dubbelriktad kommunikation mellan mag-tarmkanalen och ZNS via neurala (Vagusnerven), endokrina (HPA-axeln), immunologiska och metaboliska vägar; vid ASD dysbios, förändrad produktion av neurotransmittor-prekursorer, störda kommunikationsvägar mellan tarm och hjärna.
Hämoxygenase-1 (HO-1)
Enzym som bryter ner hem till biliverdin (antioxidativt), kolmonoxid (antiinflammatoriskt) och järn; Nrf2-reglerat; bosswellsyror ökar HO-1-uttryck; HO-1-induktion skyddar mot oxidativ stress och neuroinflammation.
Hippocampus
Hjärnstruktur i mediala tinningloben; förmedlar deklarativt minne, rumslig navigation, kontextuell bearbetning; vid ASD förstoring i barndomen, förändrad konnektivitet, störd kontextuell rädsloinlärning.
Hyperserotoninemi
Erhöhte periphere Serotonin-Spiegel; bei 25–50 % der ASD-Patienten nachgewiesen; Beziehung zu zentralen 5-HT-Spiegeln unklar; möglicherweise durch gestörte Plättchen-Serotonin-Aufnahme oder erhöhte periphere Synthese.
Interleukin-6 (IL-6)
Proinflammatoriskt cytokin; vid ASD förhöjda IL-6-nivåer i plasma och CSV; maternell immunaktivering med IL-6 producerar ASD-liknande fenotyper i musmodeller; IL-6 modulerar synaptisk transmission och utveckling.
Lillhjärnan
Hjärnstruktur i bakre hjärnan; traditionellt associerad med motorisk kontroll, spelar även roller i kognitiva processer, sensorisk prediktion och socialt lärande; vid ASD förlust av Purkinjeceller, minskad vermissvolym, störd cerebello-kortikal konnektivitet.
Lavendel (Lavandula angustifolia)
Aromatisk växt, eterisk olja innehåller 25–45 % linalool och linalylacetat; små pilotstudier rapporterar förbättrad ABA-efterlevnad och delad uppmärksamhet hos barn med ASD; prekliniska bevis för GABAerg modulering, serotonerg reglering och oxytocinaktivering.
Linalool
Monoterpenalkohol, huvudkomponent i lavendel, koriander, bergamott; modulerar GABA-A-receptorer vid bensodiazepinbindningsstället (flumazenil-känsligt), påverkar serotoninnivåer, ökar intracellulärt kalcium i oxytocinneuroner, uppvisar antiinflammatoriska effekter.
Linalylacetat
Ester av linalool och ättiksyra; huvudkomponent i lavendel (25–45 %) och bergamott; bidrar till ångestdämpande och lugnande effekter; hydrolyseras till linalool.
Långvarig potentiering (LTP) / Långvarig depression (LTD)
Cellulära mekanismer för synaptisk plasticitet; LTP: ihållande förstärkning av synaptisk transmission efter högfrekvent stimulering; LTD: ihållande försvagning efter lågfrekvent stimulering; vid ASD störd LTP/LTD-balans i multipla hjärnregioner.
Magnetresonansspektroskopi (MRS)
Icke-invasiv avbildningsteknik för kvantifiering av metaboliter (glutamat, GABA, N-acetylaspartat, kreatin) in vivo; vid ASD visar MRS regionala förändringar i glutamat- och GABA-koncentrationer.
Maternell immunaktivering (MIA)
Präklinisk modell där dräktiga djur behandlas med immunstimulerande medel (Poly(I:C), LPS); avkommor uppvisar ASD-liknande beteenden, ökat pro-inflammatoriskt cytokin, störd synaptisk utveckling; modellerar miljömässig riskfaktor för ASD.
Melatonin
Hormon som produceras av tallkottkörteln; reglerar dygnsrytm och sömn; vid ASD är sömnstörningar vanliga; melatonintillskott förbättrar insomningstid och sömnlängd; metaanalyser visar måttlig effekt.
Mikroglia
Residerande immunceller i CNS; reglerar synaptisk beskärning, neuronal utveckling, immunövervakning; vid ASD aktiverade mikroglia (ökad Iba1-expression, amöboid morfologi) i prefrontala cortex, lillhjärnan; frisätter proinflammatoriska cytokiner och ROS.
Mitokondriell dysfunktion
Nedsatt mitokondriell funktion (reducerad andningskedjeaktivitet, ökat laktat, reducerad ATP-syntes); hos 5–30 % av ASD-patienter; leder till ökad ROS-produktion, oxidativ stress, störd kalciumhomeostas.
Neuroligin (NLGN)
Postsynaptiska celladhesionsmolekyler (NLGN1-4); interagerar med presynaptiska neurexiner; reglerar synaptisk differentiering, mognad och funktion; mutationer i NLGN3 och NLGN4X hos ASD-patienter; NLGN3-R451C-möss uppvisar ökad inhibitorisk transmission och ASD-liknande fenotyper.
Neurexin (NRXN)
Presynaptiska celladhesionsmolekyler (NRXN1-3); interagerar med postsynaptiska neuroliginer; reglerar synaptisk organisation; mutationer i NRXN1 hos ASD-patienter; NRXN1-deletioner associerade med ASD, schizofreni och intellektuell funktionsnedsättning.
Neuroinflammation
Inflammatorisk reaktion i CNS; kännetecknas av mikroglia- och astrocytaaktivering, ökade proinflammatoriska cytokiner (TNF-α, IL-6, IL-1β), störd blod-hjärnbarriär; vid ASD kronisk neuroinflammation i multipla hjärnregioner.
NF-κB (kärnfaktor kappa B)
Transkriptionsfaktor som reglerar proinflammatoriska gener (TNF-α, IL-6, IL-1β, COX-2); ökad NF-κB-aktivering vid ASD; β-karyofyllen och boswelliasyror hämmar NF-κB-signalvägar.
NMDA-receptorn (N-metyl-D-aspartatreceptorn)
Jonotropisk glutamatreceptor, kalciumpermeabel kanal; central för synaptisk plasticitet (LTP/LTD), inlärning och minne; vid ASD förändrad NMDA-receptorsubunituttryck (GRIN2A, GRIN2B), störd NMDA-receptorfuntion i SHANK2-knockoutmöss.
Nrf2 (Nukleär faktor erytroid 2-relaterad faktor 2)
Transkriptionsfaktor, masterregulator av den antioxidativa responsen; inducerar expression av HO-1, NQO1, glutation-S-transferaser, andra antioxidantenzymer; boswelliasyror aktiverar Nrf2; Nrf2-aktivatorer (sulforafan) visar förbättringar vid ASD i små studier.
Oxidativ stress
Obalans mellan ROS-produktion och antioxidativt försvar; vid ASD ökad lipidperoxidation, protein-karbonylering, DNA-oxidation, reducerad antioxidantkapacitet (GSH, SOD, katalas); bidrar till nervskador och dysfunktion.
Oxytocin (OT)
Neuropeptid, producerat i hypotalamus; central roll i social bindning, tillit, social kognition, stressreglering; vid ASD minskade plasma-OT-nivåer, förändrad OXTR-uttryck; intranasal OT visar blandade resultat; OT reglerar GABAerg utveckling i prekliniska modeller.
Parvalbumin-positiva interneuron
Subtyp av GABAerg interneuron, uttrycker kalciumbindande protein parvalbumin; medierar snabb, synkroniserad inhibition; hos ASD minskad densitet och funktion av parvalbumin-interneuroner i prefrontala cortex.
Phelan-McDermid syndrom
Genetiskt syndrom orsakat av deletion på 22q13 som inkluderar genen SHANK3, karaktäriserat av autism, intellektuell funktionsnedsättning, talsvårigheter, hypotoni; demonstrerar den kausala rollen av SHANK3 i ASD-patogenes.
Prefrontala cortex (PFC)
Frontala hjärnregioner; förmedlar exekutiva funktioner, arbetsminne, kognitiv flexibilitet, social kognition, Theory of Mind; vid ASD minskad PFC-aktivering under sociala kognitionsuppgifter, störd PFC-konnektivitet, GABAerga brister.
Propionat
Kortkedjiga fettsyror, producerade av tarmmikrobiotan; vid ASD eventuellt förhöjda propionatnivåer; propionatexponering i gnagarmodeller inducerar ASD-liknande beteenden, neuroinflammation, oxidativ stress.
Reaktiva syreföreningar (ROS)
Högreaktiva molekyler (superoxid, väteperoxid, hydroxylradikal); uppstår som biprodukter av mitokondriell andning; vid överdrivna koncentrationer orsakar ROS oxidativ skada på lipider, proteiner, DNA; vid ASD ökad ROS-produktion.
Risperidon
Atypiskt antipsykotikum, dopamin-D2- och serotonin-5-HT2A-antagonist; FDA-godkänt för irritabilitet vid ASD; biverkningar: viktökning, hyperprolaktinemi, extrapiramidala symptom, metaboliskt syndrom; metaboliseras via CYP2D6.
Serotonin (5-HT, 5-Hydroxytryptamin)
Monoamin-neurotransmittor; medierar humör, ångest, sömn, aptit, social kognition; vid AST hyperserotonemi (perifer), reducerad central serotoninsyntes, genetiska varianter i 5-HTT och TPH2; Linalool och Ylang-Ylang modulerar serotonerga system.
SHANK-proteiner (SH3 och multipla ankyrinrepeat-domäner)
Postsynaptiska ställningsproteiner (SHANK1-3); organiserar glutamatreceptorer, aktincytoskelettet, signalmolekyler; mutationer i SHANK2 och SHANK3 vid ASD; SHANK3-knockoutmöss uppvisar minskad densitet av taggar, störd synaptisk transmission, ASD-liknande beteenden.
Striatum
Subkortikala strukturer i basala ganglierna (Nucleus caudatus, Putamen, Nucleus accumbens); förmedlar belöningsbearbetning, vana-inlärning, motorisk kontroll; vid ASD stratiumförstoring, minskad aktivering vid sociala belöningar, störd kortiko-striatal konnektivitet.
Synaptisk beskärning
Utvecklingsbiologiska process där överflödiga synapser elimineras; medieras av mikroglia, komplementsystemet; vid ASD möjligen störd beskärning (för mycket eller för lite), vilket leder till förändrade synapsdensiteter och konnektivitet.
Terpen
Diverse klass av naturprodukter, uppbyggda av isoprenenheter (C5H8); huvudkomponenter i eteriska oljor; monoterpener (C10): linalool, 1,8-cineol, α-pinen; sesquiterpener (C15): β-karyofyllen, cedrol; triterpener (C30): boswellinsyror.
TNF-α (Tumörnekrosfaktor-α)
Proinflammatoriskt cytokin; vid ASD ökade TNF-α-nivåer i plasma, likvor, postmortalt hjärnvävnad; maternal immunaktivering med ökad TNF-α producerade ASD-liknande fenotyper; TNF-α modulerar synaptisk transmission, minskar GABAerg hämning.
Translokatorprotein (TSPO)
Mitokondriellt protein, uppreglerat i aktiverade mikroglia; PET-ligander (t.ex. [11C]-PBR28) binder till TSPO och möjliggör in vivo-avbildning av mikroglial aktivering; vid ASD ökad TSPO-bindning i flera hjärnregioner.
Tryptofan
Essentiell aminosyra, föregångare till serotonin och melatonin; vid ASD kan tryptofanmetabolismen vara störd; tarmmikrobiotan påverkar tillgången och metabolismen av tryptofan via kynurenin- och serotoninvägarna.
Valproinsyra (VPA)
Antiepileptiskt läkemedel; prenatal VPA-exponering är en riskfaktor för ASD hos människor; VPA-inducerad ASD-musmodell: dräktiga möss får VPA, avkomman uppvisar ASD-liknande beteende, neuroinflammation, oxidativ stress; allmänt använd preklinisk modell.
Venturiprincipen
Fysikaliskt princip (Bernoullis princip): Tryckminskning i en vätska när den strömmar genom en förträngning; i Organik Aromas Nebulizer 3.0 skapar tryckluft en undertryck genom en munstycke, som suger upp eterisk olja och nebuliserar den till fina droppar (kallförstoftning).
Vetiver (Vetiveria zizanioides)
Tropiskt gräs, eterisk olja innehåller vetiverol, khusimol, α-vetivon; anekdotiska rapporter om effekter på uppmärksamhet och hyperaktivitet, men kontrollerade studier vid ASD saknas; vetenskaplig evidens för mekanismer otillräcklig.
Frankincense (Boswellia)
Harts från Boswellia-arter; eterisk olja och extrakt innehåller boswelliasyror; prekliniska bevis för anti-neuroinflammatoriska, antioxidativa (Nrf2/HO-1) och neuroprotektiva effekter; inga kliniska studier vid ASD identifierade.
Ceder (Cedrus atlantica, Juniperus virginiana)
Eterisk olja innehåller cedrol, α-cedren, thujopsen; traditionellt använd för sedering och ångestreducering, men vetenskapliga bevis för GABAerga mekanismer eller klinisk effekt vid ASD saknas.
Dygnsrytm
Endogen ~24-timmarsrytm, reglerar sömn-vakenhetscykel, hormonutsöndring, kroppstemperatur; vid AST ofta störda cirkadiska rytmer, sömnstörningar; melatonin reglerar cirkadisk rytm och förbättrar sömn vid AST.